Дрожжи занимают центральное место в исследованиях по генетике митохондрий, поскольку эти органеллы наиболее активны у таких организмов. Половой цикл одного из представителей сахаромицетов Saccharomycescerevisiaeможет быть изображён в виде такой схемы:
Представленная схема отражает существование гаплоидных и диплоидного клонов клеток с образованием гаплоидных аскоспор.
Существование внеядерной наследственности у дрожжей впервые было показано в работах Эфрусси с соавт., описавших мутацию petite. Мутантные клетки, выращенные на глюкозе, обладали пониженным дыханием и почти не содержали цитохромов а 1 ,a 3 ,b.
Генетичекий анализ показал, что мутация petiteимеет цитоплазматическую природу. Позднее было установлено, что митохондриальная ДНК цитоплазматических мутантовpetiteотличается по плавучей плотности и содержанию ГЦ-пар от ДНК митохондрий нормальных клеток этих дрожжей.
Позднее были получены цитоплазматические мутации устойчивости дрожжей к антибиотикам.
Расщепление и рекомбинация митохондриальных генов в зиготах послужили основой для построения генетических карт цитоплазматических систем.
Дальнейшие работы позволили обнаружить у дрожжей явление супрессивности (suppression– подавление), которое выражается в том, что при скрещивании дикой (нормальной) формы дрожжей с мутантной типаpetiteв зиготах происходит преимущественная репликация ДНК митохондрий мутантной формы. Через некоторое время аллель дикого типа не просто переходит в скрытое (рецессивное) или неактивное состояние, а исчезает совсем и уже никогда не проявляется у потомства.
Явление супрессивности можно рассматривать как одну из форм преимущественной передачи наследственности, при которой один родительский цитоплазматический геном берёт верх над другим. Этот процесс оказывается аналогичным материнскому наследованию у хламидомонады, о котором говорилось выше.
Позднее цитоплазматические гены были выявлены и у многих других грибов. Биохимические изменения в работе митохондрий у них также были связаны с нарушениями биосинтеза цитохромной системы.
Таким образом, описанные примеры позволили расширить арсенал экспериментальных методов для исследования цитоплазматической наследственности и показать высокую значимость цитоплазматической наследственности для обеспечения жизнедеятельности организмов.
Роль цитоплазматических генов в биогенезе клеточных органелл
С самого начала ХХ столетия цитологии постоянно подчёркивают генетическую непрерывность хлоропластов и митохондрий, а именно то, что эти органеллы возникают только из предшествующих органелл того же рода.
Изучение роли митохондриальных генов в биогенезе митохондрий показали, что там присутствуют:
специфические ферменты – РНК-полимеразы;
все три вида РНК: рибосомальные, транспортные, информационные;
особенные рибосомы.
Именно эти компоненты обеспечивают способность органелл к синтезу белка, управляемому их собственными полинуклеотидами. За счёт работы этой системы синтезируются белки только внутренней мембраны митохондрий. При этом почти все белки митохондриальных рибосом синтезируются в цитоплазме.
Изучение роли хлоропластных генов в биогенезе хлоропластов позволило выявить те же компоненты белок-синтезирующей системы, что и в митохондриях. Однако, спектр молекул белков, образующихся в хлоропластах, оказался значительно шире, что обусловлено, главным образом, значительно большей информационной ёмкостью ДНК хлоропластов. Позднее было установлено, что за счёт собственной ДНК в хлоропластах синтезируется большая субъединица центрального фермента ассимиляции СО 2 – рибулозобисфосфат-карбоксилазы, фермента фосфорибулокиназы, некоторые белковые компоненты фотосистемыII, а также белки, участвующие в построении внутренней хлоропластной мембраны, в т.ч. мембран тилакоидов, и т.д.
Дожжевая клетка имеет все основные структуры, которые присущи любой эукариотической клетке, но в то же время она обладает особенностями, свойственными грибам, а именно, сочетанием признаков как растительной, так и животной клеток: клеточная стенка у них ригидная, как у растений, но в клетке отсутствуют хлоропласты и накапливается гликоген, как у животных.
Компоненты дрожжевой клетки
Ядро
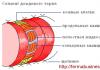
В дрожжевой клетке в фазе между делениями всегда имеется только одно ядро. В световом микроскопе его можно увидеть после специальной окраски или с помощью фазово-контрастного устройства при высоких разрешениях. На электронно-микроскопических снимках ультратонких срезов дрожжевых клеток ядро выглядит как более или менее округлая органелла, окруженная двойной мембраной. В ней есть поры в виде округлых сквозных отверстий, которые образуются в результате слияния двух ядерных мембран. Однако ядерные поры - не просто отверстия, они заполнены сложноорганизованными структурами, которые называют комплексом пор ядра. Считается, что основная функция ядерных пор - транспорт готовых рибосомных субъединиц в цитоплазму. Ядерная оболочка многофункциональна, но в основном играет роль барьера, отделяющего содержимое ядра и регулирующего транспорт макромолекул между ядром и цитоплазмой. Основные функциональные единицы ядра - молекулы ДНК, несущие основную генетическую информацию о клетке. ДНК составляет основную часть хроматина - основного компонента ядра. Число хромосом в ядре разных видов дрожжей может быть различным, оно колеблется от 2 до 16.
Митохондрии
В митохондриях имеется собственная митохондриальная ДНК (мДНК), а также весь аппарат белкового синтеза, включая матричную РНК и 70S рибосомы (в отличие от 80S рибосом в цитоплазме). мДНК у дрожжей составляет 5-20% от всей ДНК клетки. Число митохондрий в одной дрожжевой клетке варьирует в пределах 1-20 в разные периоды роста и в зависимости от условий. Как правило, 1-2 митохондрии в клетке более крупные, чем остальные и имеют разветвленную форму. Реконструкция ультратонких срезов клетки позволяет предположить, что в некоторых случаях (в подготовительный период почкования) клетка содержит всего одну вытянутую и сильно разветвленную митохондрию. Митохондрии способны к самовоспроизведению.
Цитоплазаматическая мембрана
На поперечном срезе под электронным микроскопом мембрана у дрожжей выглядит как трехслойная структура. Она представляет собой два слоя фосфолипидов, в которые погружены белковые молекулы, то есть построена по общему для всех клеточных мембран принципу. Однако, имеются различия, касающиеся химического состава. У Saccharomyces cerevisiae основными фосфолипидами мембран являются лецитин, фосфатидилэтаноламин и фосфатидилсерин. На их долю приходится около 90% всех липидов мембраны. В состав мембраны дрожжей входят стероиды - эргостерол, зимостерол и др. Белки представлены в основном ферментами, которые участвуют в трансмембранном переносе веществ, расщеплении полисахаридов и синтезе внеклеточных структур. . Функции цитоплазматической мембраны многообразны: регуляция биосинтеза клеточной стенки, активный транспорт транспорт в клетку специфических молекул органических веществ, транспорт ионов K + и Na + и др.
Вакуоли
В фазово-контрастном микроскопе в клетках дрожжей хорошо видны светлые и прозрачные структуры круглой формы. Это вакуоли. Обычно их 1-3 в клетке. Каждая вакуоль окружена одинарной мембраной и содержит различные ферменты, липиды, низкомолекулярные продукты метаболизма (аминокислоты), ионы металлов. В вакуолях сосредоточена большая часть ионов калия.
Иногда в вакуоли видны «пляшущие» за счет броуновского движения плотные гранулы. Это так называемые метахроматические гранулы, «пляшущие тельца» (dancing bodies), или волютин. Гранулы эти состоят из полимеризованных остатков фосфатов, а по периферии они покрыты комплексными соединениями из РНК, белков и липидов.
Волютин - это резерв полифосфатов в клетке. Основная функция вакуолей - разобщение процессов синтеза и распада белков и нуклеиновых кислот. Они выполняют также роль депо для хранения некоторых запасных веществ и ферментов, участвуют в регуляции тургорного давления. Также в клетке присутствуют: клеточная стенка, которая защищает протопласт от осмотического разрыва и придает клетке определенную форму; капсула (слизистый полисахаридный чехол вокруг клетки), цитоплазма и липиды.
Дожжевая клетка имеет все основные структуры, которые присущи любой эукариотической клетке, но в то же время она обладает особенностями, свойственными грибам, а именно, сочетанием признаков как растительной, так и животной клеток: клеточная стенка у них ригидная, как у растений, но в клетке отсутствуют хлоропласты и накапливается гликоген, как у животных.
Компоненты дрожжевой клетки
Ядро
В дрожжевой клетке в фазе между делениями всегда имеется только одно ядро. В световом микроскопе его можно увидеть после специальной окраски или с помощью фазово-контрастного устройства при высоких разрешениях. На электронно-микроскопических снимках ультратонких срезов дрожжевых клеток ядро выглядит как более или менее округлая органелла, окруженная двойной мембраной. В ней есть поры в виде округлых сквозных отверстий, которые образуются в результате слияния двух ядерных мембран. Однако ядерные поры - не просто отверстия, они заполнены сложноорганизованными структурами, которые называют комплексом пор ядра. Считается, что основная функция ядерных пор - транспорт готовых рибосомных субъединиц в цитоплазму. Ядерная оболочка многофункциональна, но в основном играет роль барьера, отделяющего содержимое ядра и регулирующего транспорт макромолекул между ядром и цитоплазмой. Основные функциональные единицы ядра - молекулы ДНК, несущие основную генетическую информацию о клетке. ДНК составляет основную часть хроматина - основного компонента ядра. Число хромосом в ядре разных видов дрожжей может быть различным, оно колеблется от 2 до 16.
Митохондрии
В митохондриях имеется собственная митохондриальная ДНК (мДНК), а также весь аппарат белкового синтеза, включая матричную РНК и 70S рибосомы (в отличие от 80S рибосом в цитоплазме). мДНК у дрожжей составляет 5-20% от всей ДНК клетки. Число митохондрий в одной дрожжевой клетке варьирует в пределах 1-20 в разные периоды роста и в зависимости от условий. Как правило, 1-2 митохондрии в клетке более крупные, чем остальные и имеют разветвленную форму. Реконструкция ультратонких срезов клетки позволяет предположить, что в некоторых случаях (в подготовительный период почкования) клетка содержит всего одну вытянутую и сильно разветвленную митохондрию. Митохондрии способны к самовоспроизведению.
Цитоплазаматическая мембрана
На поперечном срезе под электронным микроскопом мембрана у дрожжей выглядит как трехслойная структура. Она представляет собой два слоя фосфолипидов, в которые погружены белковые молекулы, то есть построена по общему для всех клеточных мембран принципу. Однако, имеются различия, касающиеся химического состава. У Saccharomyces cerevisiae основными фосфолипидами мембран являются лецитин, фосфатидилэтаноламин и фосфатидилсерин. На их долю приходится около 90% всех липидов мембраны. В состав мембраны дрожжей входят стероиды - эргостерол, зимостерол и др. Белки представлены в основном ферментами, которые участвуют в трансмембранном переносе веществ, расщеплении полисахаридов и синтезе внеклеточных структур. . Функции цитоплазматической мембраны многообразны: регуляция биосинтеза клеточной стенки, активный транспорт транспорт в клетку специфических молекул органических веществ, транспорт ионов K + и Na + и др.
Вакуоли
В фазово-контрастном микроскопе в клетках дрожжей хорошо видны светлые и прозрачные структуры круглой формы. Это вакуоли (см. приложение 3). Обычно их 1-3 в клетке. Каждая вакуоль окружена одинарной мембраной и содержит различные ферменты, липиды, низкомолекулярные продукты метаболизма (аминокислоты), ионы металлов. В вакуолях сосредоточена большая часть ионов калия. Иногда в вакуоли видны «пляшущие» за счет броуновского движения плотные гранулы. Это так называемые метахроматические гранулы, «пляшущие тельца» (dancing bodies), или волютин. Гранулы эти состоят из полимеризованных остатков фосфатов, а по периферии они покрыты комплексными соединениями из РНК, белков и липидов. Волютин - это резерв полифосфатов в клетке. Основная функция вакуолей - разобщение процессов синтеза и распада белков и нуклеиновых кислот. Они выполняют также роль депо для хранения некоторых запасных веществ и ферментов, участвуют в регуляции тургорного давления. Также в клетке присутствуют: клеточная стенка, которая защищает протопласт от осмотического разрыва и придает клетке определенную форму; капсула (слизистый полисахаридный чехол вокруг клетки), цитоплазма и липиды.
ДРОЖЖИ - одноклеточные грибки (грибы по бот. клас.), вегетативное размножение которых осуществляется отпочковыванием дочерних клеток, почкованием, сходным с делением, или настоящим делением. Некоторые Д. могут образовывать также разные формы мицелия, т. е. обладают дрожжемицелиальным диморфизмом. Такие Д. иногда выделяют в подгруппу дрожжеподобных грибков.
Наиболее известны Д., вызывающие брожение (см.). Большинство видов Д.- сапрофиты; немногие виды патогенны для человека, животных или растений; они вызывают дрожжевые микозы (см.).
Д. используют в хлебопечении, виноделии, пивоварении, производстве молочнокислых продуктов, спирта, глицерина и т. д.
Термин «дрожжи» охватывает филогенетически гетерогенную группу организмов, к-рую подразделяют на четыре подгруппы: 1) дрожжи - примитивные аскомицеты (см.), входящие в состав порядка Endomycetales; 2) дрожжи-базидиомицеты, принадлежащие к порядку Ustilaginales, представленные родами Rhodosporidium и Leucosporidium; 3) дрожжеподобные грибки, формирующие баллистоспоры, объединенные в семейство Sporobolomycetaceae; 4) дрожжи, у которых не обнаружено ни полового цикла, ни образования баллистоспор, отнесенные к дейтеромицетам. Четвертая подгруппа наиболее гетерогенна. Ее представителей по мере изучения переводят в первые три подгруппы в качестве новых родов и видов.
Большинство клеток Д. имеет размеры 2-3 мкм, но у некоторых видов они достигают 20-50 мкм. По форме они могут быть шаровидными, овальными, цилиндрическими, булавовидными, угловатыми, бутылковидными, веретенообразными, серповидными.
У шаровидных клеток почки формируются на любом участке поверхности, у остальных типов клеток они локализованы ближе к полюсам.
После 5-15 почкований материнская клетка гибнет. У большинства Д. клетки отделяют последовательно по одной почке, но у некоторых (Debaryomyces, Pichia) наблюдается одновременное наличие нескольких почек. При замедленном отделении почек от материнской клетки образуются цепочки из многих клеток, которые классифицируют как примитивный псевдомицелий. Истинный мицелий у Д. формируется, как у всех грибов, и состоит из гиф, разграниченных перегородками - септами. Почки псевдомицелия, образующиеся на концах разветвлений или сбоку на перетяжках, называются бластоспорами. Псевдомицелий и истинный мицелий классифицируют по характеру ветвления, обилию и размерам бластоспор, а также по формированию артроспор, к к-рым относят клетки, образующиеся при распаде нитей мицелия. Яйцевидные или эллипсоидные артроспоры называют оидиоспорами, или опциями. У некоторых Д. (Candida albicans) на псевдомицелии образуются хламидоспоры - крупные шаровидные клетки с плотной двойной стенкой. Перечисленные виды вегетативных спор встречаются у дейтеромицетов и у Д. с выраженным половым циклом. Клетки спороболомицетов образуют заостренные выросты - стеригмы, несущие на кончиках палочковидно изогнутые или симметричные баллистоспоры. При созревании эти споры с силой отбрасываются на расстояние до нескольких миллиметров.
Клетки типичных Д.- сахаромицетов имеют плотную многослойную оболочку (рис. 1). Наружный слой - гладкая липопротеидная мембрана, под ней фибриллярный слой, состоящий из комплекса маннанов с белками. Следующий слой состоит в основном из глюкана, содержащего 94% глюкозы и 6% гексозамина. Между этим слоем и цитоплазматической мембраной выделяют тонкий белковый слой. В состав оболочки входит также хитин (1 - 2%), локализованный в виде гранул. Он имеет нек-рое сходство с хитинами насекомых и иглокожих. Цитоплазматическая мембрана трехслойная, состоит из липопротеидов и образует глубокие инвагинации. Часть мембран ретикулума связана с 80 S рибосомами и входит в состав полисом. С ретикулумом связана также система комплекса Гольджи - параллельно уложенные стопки мембран - диктиосомы, по периферии образующие валик с пузырьками, которых тем больше, чем активнее процессы брожения. Производными комплекса Гольджи являются различные лизосомы: истинные лизосомы, сегрегационные гранулы, центральная вакуоль, обладающая протеолитической активностью, и звездчатые фаголизосомы, которые могут передвигаться и фагоцитировать поврежденные органоиды. Прочие многочисленные вакуоли формируются ретикулумом либо комплексом Гольджи. Некоторые депонируют волютин и липиды, другие изолируют продукты распада. Митохондрии сходны по своей структуре с митохондриями простейших. Их особенностью является большая лабильность обмена, высокая проницаемость мембран, относительно высокое содержание ДНК (1,1-4,3 мкг/мг белка) и 70 S рибосомы, аналогичные бактериальным. Ядро диам. 1 - 2 мкм содержит плотную Фейльген-положительную кариосому. Примерно половина ядра заполнена РНК, к-рой больше, чем ядерной ДНК. Из-за обилия РНК хромосомы плохо различимы. В некоторых ядрах видно веретено в виде пучка трубочек. Расхождение хромосом происходит без резорбции ядерной мембраны. После завершения митотического цикла ядро делится перетяжкой. Дочернее ядро мигрирует в почку. После отделения почки на поверхности материнской клетки остается рубец. Клетки многих Д. окружены капсулой из растворимых фосфоманнанов (Hansenula, Pichia, Pashysolen), полисахаридов (Cryptococcus, Lipomyces, Rhodotorula) или сфинголипидов (некоторые виды Hansenula).
У дрожжеподобных грибков смена типа роста сопровождается изменением состава клеток: у мицелиальных форм больше углеводов, триглицеридов, свободных жирных к-т, меньше стеролов; у дрожжевых форм повышена активность редуктазы дисульфидных связей, что приводит к изменению баланса SH-групп в составе клеточной стенки.
Д. могут существовать в гаплоидном и диплоидном состоянии. Встречаются штаммы с большей или промежуточной степенью плоидности, но это обычно сопровождается нарушением свойственного данному виду жизненного цикла. Можно выделить пять основных жизненных циклов Д. 1. Асексуальный - мейоз не обнаружен, могут образовываться вегетативные споры. Характерен для дейтеромицетов. 2. Гаплоидный - вегетативная фаза гаплоидная, образование зиготы ведет к немедленному мейозу (см.) и споруляции. Споры гаплоидные, при их прорастании возникают гаплоидные клетки. Встречаются у шизосахаромицетов и у Saccharomyces elegans. 3. Гаплоидный с дикарионом (клетка с двумя ядрами) - напоминает предыдущий цикл, но перед слиянием ядер существует стадия дикариона, к-рая может иметь разную продолжительность. Классический тип встречается у Rhodosporidium и Leucosporidium, видоизменения - у некоторых аскомицетов. 4. Гаплоидно-диплоидный - клетки в обеих фазах могут размножаться вегетативно; диплоидизация происходит либо при слиянии клеток противоположных типов спаривания (гетероталличные культуры), либо при изменении условий роста. Встречается у некоторых аскоспоровых Д. 5. Диплоидный - в гаплоидной фазе находятся только аскоспоры и в некоторых случаях первые поколения клеток после прорастания спор; затем происходит диплоидизация. Диплоидизация может происходить путем слияния дочерних ядер в прорастающей споре, слияния прорастающих спор, слияния клеток противоположного типа спаривания или слияния гаплоидной клетки и споры. В трех последних случаях за плазмогамией следует слияние ядер. У Д. этого типа гибридизация легко осуществляется смешиванием аскоспор у гомоталличных штаммов (без выраженной половой дифференциации) , вегетативных клеток противоположных типов спаривания у гетероталличных штаммов (с половой дифференциацией) или вегетативных клеток со спорами при гибридизации гетероталличного штамма с гомоталличным. У Д. родов Saccharomyces и Hansenula возможна внутри- и межвидовая гибридизация. В отдельных случаях выделены межродовые гибриды. У сахаромицетов можно получить полиплоидные гибриды.
Гаплоидизация у Д. сопровождается образованием мейотических (половых) спор. У дрожжей-аскомицетов споры заключены в аске (сумке). Их форма, величина и количество являются видовым признаком.

В ряде случаев половые споры формируются в строго определенных условиях окружающей среды, что затрудняет установление жизненных циклов и приводит к накоплению множества родовых и видовых синонимов. Для определения вида Д. необходимо получить морфол., репродуктивные и некоторые физиол, характеристики, к к-рым относятся форма вегетативных клеток, наличие и тип мицелия, форма спор и асков (рис. 2), сбраживание сахаров, ассимиляция разнообразных источников углерода и азота, осмо- и антибиотикоустойчивость, температурный максимум роста, потребность в витаминах, расщепление жиров, образование органических к-т.
Для роста Д. более благоприятны кислые субстраты, поэтому для их выделения используют среды с pH 4,6 ис 1% глюкозы. В среду часто добавляют противобактериальные антибиотики. Оптимальная температура выращивания 25-30°.
В природе Д. встречаются практически везде, но большинство видов имеет определенные места обитания: на ягодах и фруктах встречаются сахаромицеты, в морской воде, рассолах и копченых продуктах - дебариомицеты, которые могут расти при содержании NaCl до 24%; на субстратах с высоким содержанием сахара (мед, варенье) развиваются некоторые виды Zygosaccharomyces. Debaryomyces formicarius обитает только в муравейниках. Все виды Lipomyces выделяются лишь из почв. Дейтеромицеты (Candida, Trichosporon) часто обнаруживают на коже и слизистых оболочках здоровых людей. Штаммы, выделенные у людей, условно патогенны.
Среди заболеваний, вызываемых Д., наиболее часто встречаются кандидозы (см.) - поражение кожи, ногтей, слизистой оболочки ротовой полости, глотки, бронхов, легких, жел.-киш. тракта, гениталий или септическая форма. Возбудители (в порядке уменьшения встречаемости патогенных штаммов): Candida albicans, С. stellatoidea, C.tropicalis, C.pseudotropicalis, C.parapsilosis, C. guilliermondii. Эти Д. в тканях и выделениях обнаружены в виде почкующихся клеток или септированного мицелия. В культуре Candida albicans образуют хламидоспоры, в отличие от остальных потенциально патогенных видов.
Одним из возбудителей геотрихоза (см. Плесневые микозы) - заболевания слизистых оболочек - является Trichosporon cutaneum (син. Geotrichum cutaneum). Он образует прямоугольные клетки и септированный мицелий с артроспорами. Штаммы этого вида вызывают поражение толстой кишки и легких. Возбудитель криптококкоза (см.) Cryptococcus neoformans (син. Filobasidiella neoformans) поражает ц. н. с. и различные органы. Его вирулентность связана с полисахаридной капсулой, отсутствующей у невирулентных штаммов. Из споровых Д. известен лишь вид Debaryomyces klockeri (син. Deb. hansenii), выделяемый иногда при североамериканском бластомикозе (см.), однако не выяснено, патогенен ли этот вид или только сопутствует основному возбудителю Blastomyces dermatitidis. Дрожжевые микозы часто развиваются при авитаминозах, употреблении антибиотиков, подавляющих нормальную бактериальную микрофлору, применении кортикостероидов и цитостатических препаратов.

Для выделения патогенных Д. используют среду Сабуро, картофельный, кукурузный и кровяной агар, на которых Д. формируют разнообразные колонии (рис. 3). Для наблюдения клеток Д. весьма удобны прижизненная фазово-контрастная и люминесцентная микроскопия, а также витальная окраска нейтральротом. Дрожжи родов Candida и Trichosporon определяют в нативных препаратах, капсула возбудителя криптококкоза хорошо видна в капле туши. Мазки окрашивают по Граму, все Д. грамположительны. Определяют Д. по таксономическому справочнику Лоддер (J. Lodder, 1970). Приемы выделения патогенных Д. описаны Н. П. Ели-новым.
Д., помимо основного использования В хлебопекарной и пивоваренной промышленности, могут применяться как ценный питательный продукт, дополнительный источник высокоценного белка (см. Белки одноклеточных), витаминов группы В и D и минеральных веществ. Д. могут сыграть важную роль в повышении уровня белка в питании населения многих стран мира, испытывающих выраженную белковую недостаточность.
Особую ценность и перспективность в питании человека имеет чистый белок - изолят, получаемый из Д. Будучи освобожден от всех нежелательных и небезразличных для организма веществ (нуклеиновых к-т, пуринов и др.), изолят Д. может широко использоваться для производства белковых концентратов, искусственных пищевых продуктов, а также для обогащения белком натуральных пищевых продуктов (хлеба, муки, крупы и др.).
Хим. состав Д. зависит от вида и расы грибка, состава питательной среды, условий культивирования и ряда других факторов.
Сухие пищевые Д. содержат 50% белка, 25-40% углеводов и до 3% жира.
Все виды Д. отличаются высоким содержанием азотистых веществ. Содержание азота в сухих Д. составляет в среднем 7-9% (на сухой вес). Из этого количества 18-20% приходится на небелковый азот, при этом 8-13% составляют пуриновые основания, ок. 4% - пиримидиновые основания, 0,5% - глюкозамин и др.
Белок Д. характеризуется сбалансированностью аминокислот, близкой к животному белку, за исключением содержания метионина, которого в 2-3 раза меньше, чем в белке мяса и других животных продуктов (табл.).
Таблица. Содержание незаменимых аминокислот в некоторых видах продуктов из дрожжей
|
Наименование продукта |
Аминокислоты в процентах к общему азоту |
||||||
|
триптофан |
метионин |
изолейцин |
|||||
|
Снятое молоко |
|||||||
|
Говядина |
|||||||
|
Дрожжи «углеводные» |
|||||||
|
Дрожжи «нефтяные» |
|||||||
|
Дрожжи «газовые» |
|||||||
Примечание. К «углеводным» относятся все виды Д. пищевого предназначения; «нефтяные» и «газовые» Д., находящиеся в стадии изучения, относятся к кормовым Д.
Несмотря на сравнительно невысокое содержание метионина, белок Д. является высокоценным; в смешанной диете в сочетании с белками других продуктов, напр, мяса и молока, он может служить существенным источником лабильных метильных групп, обеспечивающих синтез холина в организме. Белок Д. хорошо переваривается и усваивается. Так, перевариваемость пекарских Д. составляет ок. 80%, пивных - 80-91%, кормовых - 85-88%. Перевариваемость сухих Д. в организме человека 70-90%.
Д. относятся к наиболее богатым источникам витаминов группы В, витамина D. Они содержат эргостерол, способный при облучении УФ-лучами образовать ряд кристаллических веществ, в т. ч. и кальциферол (витамин D2). Содержание эргостерола в Д. составляет 0,6% (на сухое вещество). Д. широко используются витаминной промышленностью для производства препаратов витамина D.
В состав жира Д. входят преимущественно пальмитиновая (75%) и стеариновая (25%) к-ты, фосфатиды, а также эргостерин, количество к-рого в некоторых видах Д. достигает 2% к общему количеству жира.
В составе фосфатидов Д. находятся (в процентах к общему количеству фосфатидов) фосфатидилсерин - 26%, кефалин - 31%, нейтральные жиры - ок. 30%, мезокефалии - 13%, сфингомиелин - 1%.
Количество минеральных солей в Д. колеблется в зависимости от условий их культивирования; содержание их в процентах солеобразующих окислов следующее: K 2 O - 29,33-39,5; Na 2 O - 0,5-2,26; CaO - 1,0-7,5; MgO - 3,77-6,34; Fe 2 O 3 - 0,06- 0,7; P 2 O 5 - 44,8-59,4; SO 3 - 0,57- 6,38; SiO 2 - 0,92-1,88.
Д. используют для приготовления кулинарных и хлебобулочных изделий (см. Хлеб, хлебопродукты). Для повышения их питательной ценности можно добавлять Д. в количествах, значительно превышающих нормы хлебопечения. Для усиления белкового и витаминного питания рекомендуются сухие Д.- 10-25 г в сутки. Увеличивать эти нормы нецелесообразно, т. к. Д. содержат относительно много пурина и систематическое потребление их в большом количестве (особенно в пожилом возрасте) может отрицательно сказаться на состоянии организма.
При использовании Д. в питании необходимо учитывать возможность присутствия в них ряда нежелательных веществ, в т. ч. малоизвестных и недостаточно изученных. К ним относятся необычные компоненты липидной фракции (стерины, ненасыщенные жирные к-ты и др.), некоторые аминокислоты, в т. ч. их D-оптические изомеры аминосахаров и пептидов, особенно в составе клеточных мембран, а также ряд других биологически активных веществ известной и неизвестной природы. Только одно разрушение клеток Д. уже приводит к улучшению качества продукта и повышению усвояемости белка Д. Обработка Д. в среде органического растворителя освобождает их от липидных и других нежелательных компонентов. Отделению подлежат также нуклеиновые к-ты.
При использовании Д. в питании их нужно инактивировать путем интенсивной тепловой обработки (проваривание, прожаривание). Инактивированные Д. добавляют в различные блюда - овощные и крупяные первые блюда, в запеканки, оладьи, котлеты, жаркое, соуса и др. Для пищевых целей используют Д. прессованные, сухие и жидкие.
Для мед. целей наибольшее практическое применение получили пивные Д., как свежие - жидкие, так и сухие. Согласно стандарту (ГОСТ 171 - 69) влажность прессованных Д. должна быть не более 75%, кислотность (в пересчете на уксусную к-ту, в мг): в день выпуска Д. с завода - не более 120, а после 12 сут. хранения или транспортировки при t° 0-4° - не более 360 на 100 г д.
Сухие Д. получают путем обезвоживания прессованных Д. Сухие мед. пивные Д. поступают в торговую сеть в виде порошка. По органолептическим свойствам они должны иметь специфический дрожжевой запах без посторонних примесей, без признаков плесени и разложения органических веществ. Влажность их не должна превышать 10%, количество белковых веществ - не ниже 50% и золы - не более 10% на сухое вещество. Используя современные технологические методы производства, можно получить высокоактивные сухие Д., сохраняющие активность в течение длительного срока.
Жидкие Д.- крайне нестойкий продукт, подлежащий реализации в течение первых 6-8 час. Для удлинения срока хранения и возможности транспортировки жидкие Д. можно подвергать специальной обработке путем уваривания и добавления соли.
Д., особенно прессованные и жидкие, относятся к скоропортящимся продуктам, требующим строгого ограничения сроков хранения при t° 0-4°. Прессованные Д. допускается хранить при t° 0-4° в течение 10-12 сут., жидкие и питьевые - 6-8 час., дрожжевую пасту - ок. 3 сут.
Д. находят применение в некоторых отраслях пищевой промышленности. В кондитерской и сахарной промышленности Д. могут использоваться как источник инвертазы- фермента, осуществляющего инверсию сахарозы.
Особое значение имеют Д. для повышения пищевой ценности галет и печенья путем обогащения их дрожжевым белком и группой витаминов В.
Галеты, обогащенные Д., содержат в 1 кг: белка - 105 г, жира - 70 г, витамина B1 - 9-12 мг, B2 - 7-9 мг, PP - 90-95 мг, фолиевой к-ты - 75-80 мг, холина - 4300 мг и другие витамины группы В. Кроме того, белок таких галет содержит повышенный комплекс незаменимых аминокислот.
Исследование Д. производят путем определения органолептических показателей и лабораторного анализа - согласно методике, приведенной в соответствующих стандартах (ГОСТ 171 - 69).
Большое значение имеет производство Д., источником энергии которых служат нормальные парафины и углеводороды.
Это открывает большие возможности для применения углеводородных Д. в качестве важного источника кормовых ресурсов и в перспективе для питания человека.
Лечебное применение дрожжей
Пищевые Д.- весьма ценный продукт для здорового и больного человека. Они имеют важное значение в профилактике алиментарной дистрофии и многих заболеваний, связанных с белковой недостаточностью.
Д. нашли широкое применение при лечении больных алиментарной дистрофией, выздоравливающих после тяжелых заболеваний (острые инфекции, ожоги, травмы), больных, страдающих хрон, заболеваниями (туберкулез, бруцеллез, ревматизм и др.), а также больных пеллагрой, спру, бери-бери, анемией, фурункулезом и др. При этом преследуется цель повышенного введения в организм с рационом полноценного белка, витаминов, особенно тиамина и рибофлавина.
Дозировка Д. может быть различной в зависимости от состояния здоровья больных и от степени белковой и витаминной недостаточности. Суточная доза для взрослых 50 г прессованных или 25 г сухих. Для детей младших возрастов- 25-30 г прессованных или 10 г сухих Д. в сутки.
Пекарские Д. вводят в первое и второе блюдо или закуски либо из Д. готовят дрожжевой напиток или специальные сорта хлеба. Сухие и прессованные Д. перед закладкой в изготовляемые блюда необходимо подвергнуть тепловой обработке.
Прессованные пищевые Д. кладут в соответствующую посуду (кастрюлю), в к-рую добавляют воду и соль из расчета 30 г воды и 15 г соли на 1 кг Д. Посуду с Д. ставят в другую посуду с горячей водой и варят под крышкой в течение 1 часа, после чего посуду с Д. ставят на горячую (но не раскаленную) плиту и варят еще 1 час при постоянном помешивании. При этом способе тепловой обработки прессованные Д. теряют присущий сырым Д. запах и вкус; происходит также разрушение дрожжевых клеток и бродильных элементов. Сухие пекарские Д. подвергают такой же тепловой обработке, но с предварительным разведением их холодной водой из расчета 4 л воды на 1 кг сухих Д. Жидкие Д. выпаривают до состояния густой пасты. Подвергшиеся тепловой обработке Д. закладывают в изготовляемые блюда и соусы. Во вторые блюда можно добавлять прожаренные Д. При высоких температурах Д. дают резкий, неприятный запах, поэтому в качестве белкового обогатителя используют комплексный дрожжевой ферментный препарат, который готовят из отмытых Д., обработанных ультразвуком. Под действием ультразвука дрожжевые клетки распадаются, а аминокислоты, ферменты, витамины и другие вещества остаются в неденатурированном виде. Комплексный дрожжевой ферментный препарат широко используют в хлебопекарной промышленности и для приготовления различных блюд.
Пивные Д., в которых содержатся безазотистые и азотистые вещества, ферменты и витамины (B 1 , B 2 , PP, а также провитамин D - эргостерин), используют для мед. целей в сухом виде при гиповитаминозе В 1 , фурункулезе, а также при различных нарушениях обмена веществ, сопровождающихся витаминной недостаточностью. Из высушенных пивных Д. готовят р-р с сахаром, содержащий не менее 3 мг% витамина B1. Выпускают этот р-р во флаконах по 0,5 л и назначают внутрь по 1 чайн. л. 2-3 раза в день взрослым и 1 - 2 раза в день детям.
С леч. целью применяют также сухие пивные Д. (содержание витамина B 1 не менее 14 мг%). Сухие пивные Д. иногда назначают вместе с фитином (таблетки гефефитина, содержащие 0,375 г сухих пивных Д. и 0,125 г фитина) по 2-3 таблетки 2-3 раза в день.
Противопоказание для применения Д.- заболевания, при которых нужно ограничивать в рационе экстрактивные вещества (заболевания почек с синдромом хрон, почечной недостаточности, подагра, хрон, сердечно-сосудистая недостаточность и др.), а также индивидуальная непереносимость Д.
Библиография: Бирюзова В.П. Мембранные структуры микроорганизмов, М., 1973, библиогр.; Генетические основы селекции микроорганизмов, под ред. С.И. Алиханяна, с. 59, М., 1969, библиогр.; E л и н о в Н. П. Патогенные дрожжеподобные микроорганизмы, М., 1964, библиогр.; P h a f f H. J., Miller M. W. a. M r a k E. M. The life of yeasts, Cambridge, 1966; The yeasts, A taxonomic study, ed. by J. Lodder, Amsterdam - L., 1970; The yeasts, ed. by A. H. Rose a. J. S. Harrison, v. 1-3, L."- N. Y., 1969-1971.
Г. Ф. Нестерова, Я. О. Соом: К. С. Петровский (пит.), М. А. Самсонов (леч. применение).
Ученые выяснили, что при почковании клетки дрожжей передают своему потомству больше митохондрий, чем оставляют себе.
Порыв пожертвовать всем, даже собственным здоровьем, на благо своего потомства присущ различным видам живых существ, а не только людям. Самки полярных медведей умирают от голода, дельфины-матери перестают спать, а некоторые виды пауков жертвуют собой, чтобы обеспечить потомство питанием.
Наука и жизнь // Иллюстрации
Наука и жизнь // Иллюстрации
Необычайное открытие сделали учёные из Университета Калифорнии, Сан-Франциско (UCSF). Оказалось, что даже дрожжам присущ «родительский инстинкт», и они могут пожертвовать собой, чтобы потомство смогло выжить. Согласно данным, опубликованным в журнале Science, специалисты из UCSF обнаружили, что пекарские дрожжи (Saccharomyces cerevisiae) передают большую часть своих митохондрий потомству. Митохондрии – миниатюрные «электростанции» клеток растений, животных и грибов (к коим относят и дрожжи), вырабатывающие энергию для протекания основных биохимических процессов.
Долгое время считалось, что при митозе – процессе деления клеток, все клеточные органеллы делятся поровну. Но так происходит не со всеми клетками. Стволовые клетки человека, например, часто делятся таким образом, что получившиеся в результате клетки и «выглядят», и «ведут себя» по-разному. То же самое происходит с некоторыми раковыми клетками. Процесс митоза у дрожжей называют почкованием. Его особенность в том, что при почковании (делении) материнской клетки потомству достается больше митохондрий, чем остается в родительской клетке. «Перекачка» дополнительных митохондрий происходит при участии белков цитоскелета. Обычно «материнского капитала» хватает на 10 делений, к 20-му практически все материнские клетки умирают. Любопытно, что сама «дочерняя» клетка при этом по размеру меньше «материнской».
Больше всего ученых удивил сам факт, что «материнские» дрожжи передают митохондрии потомству, приближая тем самым собственную смерть.
Руководитель исследования Уоллас Маршалл, доктор биохимических и биофизических наук, сообщил, что материнская клетка будет передавать столько митохондрий, сколько понадобится новым клеткам. «Мать отдаёт всё, не получая ничего по мере роста потомства», – подчеркнул он.
Если ученые научатся контролировать процесс передачи митохондрий у дрожжей, возможно, это станет еще одним шагом к пониманию того, каким образом происходит разрастание раковых клеток.
Иллюстрации: 1. Процесс почкования Saccharomyces cerevisiae. 2. Компьютерное изображение митохондриальной сети дрожжей.