Органы зрения развиты у большинства насекомых. Наибольшего развития достигают сложные, или фасеточные, глаза . Число зрительных элементов - омматидиев, или фасеток, в глазу комнатной мухи достигает 4 тыс., а у стрекоз даже 28 тыс. Омматидий состоит из прозрачного хрусталика, или роговицы, в виде двояковыпуклой линзы и лежащего под ней прозрачного хрустального конуса. Вместе они составляют оптическую систему. Под конусом расположена сетчатка, воспринимающая световые лучи. Клетки сетчатки соединены нервными волокнами с зрительными долями мозга. Каждый омматидий окружают пигментные клетки.
В зависимости от восприятия света различной интенсивности различают аппозиционный и суперпозиционный типы глаз. Первая разновидность строения глаз характерна дневным насекомым, вторая – ночным.
В аппозиционном глазе каждый омматидий изолирован в своей верхней части при помощи пигмента от соседних омматидиев. Таким образом, каждая структурная единица глаза работает отдельно от всех остальных, воспринимая только «свою» часть внешнего пространства. Общая картинка складывается в мозге насекомого как бы из множества кусочков мозаики.
В суперпозиционном глазе омматидии лишь частично, хоть и по всей длине, защищены от боковых лучей: они полупроницаемы. С одной стороны, это мешает насекомым при интенсивном освещении, с другой – помогает им лучше видеть в сумерках.
Глазки́ (дорсальные простые глаза) – это мелкие органы зрения, которые имеются у некоторых имаго и располагаются обычно на верхней части головы. Обычно представлены в количестве трех, при этом, один лежит чуть впереди, а еще два – сзади и сбоку от переднего. В их составе нет омматидия, строение простых глазков значительно упрощено. Снаружи располагается роговица, состоящая из корнеагенных клеток, глубже находится световоспринимающий аппарат из ретинальных (чувствительных) клеток, еще ниже лежат пигментные клетки, которые переходят в волокна зрительного нерва.
Из всех разновидностей глаз насекомых простые глазки обладают наиболее слабой способностью к зрению. По некоторым данным, они вообще не выполняют зрительной функции, и лишь отвечают за улучшение функции сложных глаз. Это, в частности, доказывается тем, что у насекомых практически не бывает простых глазков в отсутствии сложных. Кроме того, при закрашивании фасеточных глаз насекомые перестают ориентироваться в пространстве, даже если у них имеются хорошо выраженные простые глазки.
Стеммы, или латеральные простые глаза – имеются у личинок насекомых с полным превращением. Во время стадии куколки они «превращаются» в сложные глаза. Выполняют зрительную функцию, но, в связи с упрощенной структурой, видят относительно слабо. Для улучшения зрения личиночные глазки нередко представлены у личинок в количестве нескольких штук. У личинок пилильщиков они сходны с дорсальными, а у гусениц бабочек напоминают омматидий сложного глаза. Гусеницы воспринимают форму предметов, различают мелкие детали на их поверхности.
Функции хордотональных органов, по-видимому, различны. В тех случаях, когда сенсиллы примыкают к кутикуле, они, как правило, служат для восприятия низкочастотных вибраций. Правда, в отдельных случаях (хордотональные органы, расположенные в антеннах комаров) они чувствительны и к колебаниям высокой частоты. Внутренние хордотональные органы, вероятно, регистрируют изменения давления и механических напряжений, возникающих в теле насекомого.
Настоящими слуховыми органами насекомых являются тимпанальные органы, в которых сколпофоры связаны с тонкими кутикулярными мембранами (тимпанальными мембранами), играющими роль барабанных перепонок.
Типичным строением обладают тимпанальные органы кузнечиков, расположенные на голенях передних ног. В верхней части голени имеются по две узкие продольные щели, ведущие в два барабанных кармана. Внутренние стенки кармашков, обращенные друг к другу, тонки и представляют собой барабанные перепонки, наружные же утолщены и называются барабанными крышечками. Между обеими барабанными перепонками, вплотную примыкая к ним, проходят два трахейных ствола, которые, быть может, служат в качестве резонаторов. Наконец, главную часть тимпанального органа составляют три группы сколпофоров. Сколпофоры примыкают частью к барабанной перепонке, частью к резонирующей трахее. Центральные отростки чувствительных клеток образуют тимпанальный нерв. Точно по такому же принципу - сочетание сколпофоров и тимпанальных перепонок - устроены тимпанальные органы и других насекомых - саранчовых, сверчков, бабочек и др. Правда, располагаться они могут в разных местах тела - на передних сегментах брюшка, у основания крыльев и т. п.
Хордотональные сенсиллы тимпанальных органов служат для восприятия колебаний различной частоты - имеются "высокочастотные" и "низкочастотные" сенсиллы. Как правило, одна из таких групп настроена на частоты, максимально представленные в звуках, издаваемых особями того же вида. В целом насекомые воспринимают звуки в очень широком диапазоне: от инфразвука (8-10 Гц) до ультразвука (45000 Гц).
Насекомые способны не только воспринимать, но и издавать звуки. Эта особенность характерна для представителей многих групп: прямокрылых , жуков , перепончатокрылых , бабочек и др. Звуковые органы насекомых очень разнообразны.
Стрекотание прямокрылых, например, вызывается развитием известных стрекочущих приспособлений, которые чаще всего связаны с крыльями. Так, у кузнечиковых эти органы находятся на передних крыльях. Некоторые жилки левого крыла становятся зазубренными и превращаются в так называемый смычок, которым животное водит по правому крылу, где в соответствующем месте находится резонатор. Последний состоит из ограниченной высокой жилкой площадки на крыле - зеркальца. Движение зазубренного смычка по краю зеркальца приводит к вибрации растянутой на нем части поверхности крыла.
У саранчовых смычок образован рядом мельчайших зубчиков на бедрах задних ног. При трении бедер о верхние крылья зубчики задевают за сильно выдающуюся у самца радиальную жилку крыла. У самцов цикад есть своеобразный "голосовой аппарат" на нижней стороне заднегруди: действие его основано на чрезвычайно быстром колебании хитиновой перепонки, приводимой в движение сокращением мышц. Значение способности издавать звуки заключается, по-видимому, в привлечении стрекочущими самцами самок.
Хеморецепторы насекомых представлены обонятельными и вкусовыми сенсиллами. Кутикулярные образования обонятельных сенсилл очень разнообразны по форме: щетинки, конусовидные придатки, пластинки и т. п. Общая черта - наличие тонких пор, пронизывающих кутикулу. Через эти поры открыт доступ к чувствительным элементам сенсиллы для молекул пахучих веществ. Обонятельные сенсиллы располагаются главным образом на сяжках и челюстных щупиках.
Обоняние служит насекомым как для отыскания пищи, так и при спаривании: самцы часто находят самок по запаху. Последние выделяют особые пахучие вещества - половые аттрактанты. Достаточно ничтожного количества (100 молекул в 1 см 3 воздуха) такого вещества, чтобы вызвать возбуждение у самцов шелкопряда.
Вкусовые сенсиллы располагаются у насекомых на ротовых конечностях и дистальных члениках лапок. Их кутикулярные элементы представлены волосками или конусовидными придатками и также пронизаны порами. В состав каждой сенсиллы входит несколько рецепторных клеток, каждая из которых реагирует на определенный вкусовой раздражитель: одна клетка реагирует на соли, другая на сахаристые вещества, третья на чистую воду. Одна из чувствительных клеток вкусовой сенсиллы является механорецепторной. Таким образом, у насекомых, так же как и у позвоночных, вкусовое ощущение сопровождается осязательным.
Наиболее сложными из органов чувств у насекомых являются органы зрения . Последние представлены образованиями нескольких типов, из которых важнейшие - сложные фасетированные глаза примерно такого же строения, как и сложные глаза ракообразных.
Глаза состоят из отдельных омматидиев, количество которых определяется главным образом биологическими особенностями насекомых. Активные хищники и хорошие летуны, стрекозы обладают глазами, насчитывающими до 28 000 фасеток в каждом. В то же время муравьи (отр. Перепончатокрылые), особенно рабочие особи видов, обитающих под землей, имеют глаза, состоящие из 8-9 омматидиев.

Каждый омматидий представляет совершенную фотооптическую сенсиллу. В его состав входят оптический аппарат, включающий роговицу, - прозрачный участок кутикулы над омматидием и так называемый хрустальный конус. В совокупности они выполняют роль линзы. Воспринимающий аппарат омматидия представлен несколькими (4-12) рецепторными клетками; специализация их зашла очень далеко, о чем говорит полная утрата ими жгутиковых структур. Собственно чувствительные части клеток - рабдомеры - представляют скопления плотно упакованных микроворсинок, располагаются в центре омматидия и тесно прилегают друг к другу. В совокупности они образуют светочувствительный элемент глаза - рабдом.
По краям омматидия залегают экранирующие пигментные клетки; последние довольно существенно отличаются у дневных и ночных насекомых. В первом случае пигмент в клетке неподвижен и постоянно разделяет соседние омматидии, не пропуская световые лучи из одного глазка в другой. Во втором случае пигмент способен перемещаться в клетках и скапливаться только в их верхней части. При этом лучи света попадают на чувствительные клетки не одного, а нескольких соседних омматидиев, что заметно (почти на два порядка) повышает общую чувствительность глаза. Естественно, что подобного рода адаптация возникла у сумеречных и ночных насекомых. От чувствительных клеток омматидия отходят нервные окончания, образующие зрительный нерв.
Кроме сложных глаз многие насекомые имеют еще и простые глазки, строение которых не соответствует строению одного омматидия. Светопреломляющий аппарат линзообразной формы, сразу же под ним расположен слой чувствительных клеток. Весь глазок одет чехлом из пигментных клеток. Оптические свойства простых глазков таковы, что воспринимать изображения предметов они не могут.
Личинки насекомых в большинстве случаев обладают только простыми глазками, отличающимися, однако, по строению от простых глазков взрослых стадий. Никакой преемственности между глазками взрослых особей и личинок не существует. Во время метаморфоза глаза личинок полностью резорбируются.
Зрительные способности насекомых совершенны. Однако структурные особенности сложного глаза предопределяют особый физиологический механизм зрения. Животные, имеющие сложные глаза, обладают "мозаичным" зрением. Малые размеры омматидиев и их обособленность друг от друга приводят к тому, что каждая группа чувствительных клеток воспринимает лишь небольшой и сравнительно узкий пучок лучей. Лучи, падающие под значительным углом, поглощаются экранирующими пигментными клетками и не достигают светочувствительных элементов омматидиев. Таким образом, схематично каждый омматидии получает изображение только одной небольшой точки объекта, находящегося в поле зрения всего глаза. Вследствие этого изображение складывается из стольких световых точек, отвечающих различным частям объекта, на сколько фасеток падают перпендикулярно лучи от объекта. Общая картина комбинируется как бы из множества мелких частичных изображений путем приложения их одного к другому.
Восприятие цвета насекомыми также отличается известным своеобразием. Представители высших групп Insecta имеют цветовое зрение, основанное на восприятии трех основных цветов, смешение которых и дает все красочное многообразие окружающего нас мира. Однако у насекомых по сравнению с человеком наблюдается сильный сдвиг в коротковолновую часть спектра: они воспринимают зелено-желтые, синие и ультрафиолетовые лучи. Последние для нас невидимы. Следовательно, цветовое восприятие мира насекомыми резко отличается от нашего.
Функции простых глазков взрослых насекомых требуют еще серьезного изучения. По-видимому, они в какой-то мере "дополняют" сложные глаза, влияя на активность поведения насекомых в разных условиях освещенности. Кроме того, было показано, что простые глазки наряду со сложными глазами способны воспринимать поляризованный свет.
Помимо перечисленных органов чувств насекомые обладают еще рядом рецепторных аппаратов. Таковы сенсиллы, воспринимающие температуру окружающей среды, ее влажность. Водные насекомые способны регистрировать изменения давления и т. п.
Органы дыхания.
Для дыхания служит сложно развитая система трахей. По бокам тела находится до 10 пар, иногда меньше, дыхалец, или стигм: они лежат на средне- и заднегруди и на 8 члениках брюшка.

Стигмы часто снабжены особыми замыкательными аппаратами и ведут каждая в короткий поперечный канал, а все поперечные каналы соединены между собой парой (или больше) главных продольных трахейных стволов. От стволов берут начало более тонкие трахеи, ветвящиеся многократно и опутывающие своими разветвлениями все органы. Заканчивается каждая трахея концевой клеткой с радиально расходящимися отростками, пронизанными конечными канальцами трахеи. Концевые веточки этой клетки (трахеолы) проникают даже внутрь отдельных клеток тела. Иногда трахеи образуют местные расширения, или воздушные мешки, которые служат у наземных насекомых для улучшения вентиляции воздуха в трахейной системе, а у водных, вероятно, в качестве резервуаров, увеличивающих запас воздуха в теле животного. Трахеи возникают у зародыша насекомых в виде глубоких впячиваний эктодермы; как и остальные эктодермальные образования, они выстланы кутикулой. В поверхностном слое последней образуется спиральное утолщение, придающее трахее эластичность и препятствующее спадению стенок.
В простейших случаях поступление кислорода в трахейную систему и удаление из нее углекислого газа происходит путем диффузии через постоянно открытые стигмы. Это наблюдается, однако, только у малоактивных насекомых, обитающих в условиях повышенной влажности.
Насекомые в настоящее время являются самой процветающей группой животных на Земле.
Тело насекомых подразделяется на три отдела: голову, грудь и брюшко.
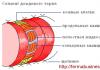
На голове насекомых находятся фасеточные глаза и четыре пары придатков. У некоторых видов кроме фасеточных глаз имеются простые глазки. Первая пара придатков представлена усиками (антеннами), являющимися органами обоняния. Остальные три пары образуют ротовой аппарат. Верхняя губа (лабрум), непарная складка, прикрывает верхние челюсти. Вторая пара ротовых придатков образует верхние челюсти (мандибулы), третья пара - нижние челюсти (максиллы), четвертая пара срастается и образует нижнюю губу (лабиум). На нижней челюсти и нижней губе могут находиться по паре щупиков. К ротовому аппарату относится язык (гипофаринкс) - хитиновое выпячивание дна ротовой полости (рис. 3). В связи со способом питания, ротовые аппараты могут быть различных типов. Различают грызущий, грызуще-лижущий, колюще-сосущий, сосущий и лижущий типы ротовых аппаратов. Первичным типом ротового аппарата следует считать грызущий (рис. 1).
рис. 1.
1 - верхняя губа, 2 - верхние челюсти, 3 - нижние челюсти, 4 - нижняя губа,
5 - основной членик нижней губы, 6 - "стволик" нижней губы, 7 - нижнечелюстной щупик,
8 - внутренняя жевательная лопасть нижней челюсти, 9 - наружная
жевательная лопасть нижней челюсти, 10 - подподбородок,
11 - ложный подбородок, 12 - нежнегубной щупик, 13 - язычок, 14 - придаточный язычок.
Грудь состоит из трех сегментов, которые называются соответственно переднегрудь, среднегрудь и заднегрудь. Каждый из сегментов груди несет пару конечностей, у летающих видов на средне- и заднегруди находятся по паре крыльев. Конечности членистые. Основной членик ноги называется тазик, за ним следуют вертлуг, бедро, голень и лапка (рис. 2). В связи с образом жизни конечности бывают ходильными, бегательными, прыгательными, плавательными, копательными и хватательными.

рис. 2. Схема строения
конечности насекомого:
1 - крыло, 2 - тазик, 3 - вертлуг,
4 - бедро, 5 - голень, 6 - лапка.

рис. 3.
1 - фасеточные глаза, 2 - простые глазки, 3 - мозг, 4 - слюнная
железа, 5 - зоб, 6 - перднее крыло, 7 - заднее крыло, 8 - яичник,
9 - сердце, 10 - задняя кишка, 11 - хвостовая щетинка (церка),
12 - антенна, 13 - верхняя губа, 14 - мандибулы (верхние
челюсти), 15 - максиллы (нижние челюсти), 16 - нижняя губа,
17 - подглоточный ганглий, 18 - брюшная нервная цепочка,
19 - средняя кишка, 20 - мальпигиевы сосуды.
Число сегментов брюшка варьирует от 11 до 4. На брюшке у низших насекомых имеются парные конечности, у высших насекомых они видоизменяются в яйцеклад или другие органы.
Покровы представлены хитиновой кутикулой, гиподермой и базальной мембраной, защищают насекомых от механических повреждений, потери воды, являются наружным скелетом. Насекомые имеют много желез гиподермального происхождения: слюнные, пахучие, ядовитые, паутинные, восковые и пр. Окраска покровов насекомых обуславливается пигментами, содержащимися в кутикуле или гиподерме.

рис. 4. Продольный разрез через
голову черного таракана:
1 - ротовое отверстие, 2 - глотка,
3 - пищевод, 4 - головной мозг
(надглоточный нервный узел),
5 - подглоточный нервный узел,
6 - аорта, 7 - проток слюнных
желез, 8 - гипофаринкс, или
подглоточник, 9 - предротовая
полость, 10 - передний отдел
предротовой полости, или
цибарий, 11 - задний отдел
предротовой полости,
или саливарий.
Мышцы насекомых по гистологическому строению относятся к поперечнополосатым, они отличаются способностью к очень высокой частоте сокращений (до 1000 раз в секунду).
Пищеварительная система как у всех членистоногих подразделяется на три отдела, передний и задний отделы имеют эктодермальное происхождение, средний - энтодермальное (рис. 5). Пищеварительная система начинается ротовыми придатками и ротовой полостью, в которую открываются протоки 1-2 пар слюнных желез. Первая пара слюнных желез вырабатывает пищеварительные ферменты. Вторая пара слюнных желез может видоизменяться в паутинные или шелкоотделительные железы (гусеницы многих видов бабочек). Протоки каждой пары соединяются в непарный канал, который открывается у основания нижней губы под гипофаринксом. Передний отдел включает в себя глотку, пищевод и желудок. У некоторых видов насекомых пищевод имеет расширение - зоб. У видов, питающихся растительной пищей, в желудке находятся хитиновые складки, зубцы, способствующие перетиранию пищи. Средний отдел представлен средней кишкой, в которой происходит переваривание и всасывание пищи. В своей начальной части средняя кишка может иметь слепые выросты (пилорические придатки). Пилорические придатки функционируют как пищеварительные железы. У многих насекомых, питающихся древесиной, в кишечнике поселяются симбиотические простейшие и бактерии, выделяющие фермент целлюлазу и этим способствующие перевариванию клетчатки. Задний отдел представлен задней кишкой. На границе между средней и задней отделами в просвет кишечника открываются многочисленные слепо замкнутые мальпигиевы сосуды. Задняя кишка имеет ректальные железы, которые отсасывают из остатков пищевой массы воду.

рис. 5. Схема строения
пищеварительной системы
черного таракана:
1 - слюнные железы, 2 -
пищевод, 3 - зоб, 4 -
пилорические придатки,
5 - средняя кишка,
6 - мальпигиевы сосуды,
7 - задняя кишка,
8 - прямая кишка.
Органы дыхания насекомых - трахеи, по которым осуществляется транспорт газов. Трахеи начинаются отверстиями - дыхальцами (стигмами), которые находятся по бокам среднегруди и заднегруди и на каждом членике брюшка. Максимальное число дыхалец - 10 пар. Часто стигмы имеют особые замыкательные клапаны. Трахеи имеют вид тонких трубочек и пронизывают все тело насекомого (рис. 6). Концевые ветви трахей заканчиваются звездчатой трахейной клеткой, от которой отходят еще более тонкие трубочки - трахеолы. Иногда трахеи образуют небольшие расширения - воздушные мешки. Стенки трахей выстланы тонкой кутикулой, имеющие утолщения в форме колец и спиралей.
рис. 6. Схема
строения
дыхательной
системы черного
таракана
Кровеносная система насекомых - незамкнутого типа (рис. 7). Сердце находится в перикардиальном синусе на дорсальной стороне брюшного отдела тела. Сердце имеет вид трубки, слепо замкнутой на заднем конце. Сердце разделено на камеры, каждая камера имеет по бокам парные отверстия с клапанами - остии. Число камер - восемь и меньше. К каждой камере сердца подходят мышцы, обеспечивающие ее сокращение. Волна сокращений сердца от задней камеры кпередней обеспечивает одностороннее движение крови вперед.
Гемолимфа движется из сердца в единственный сосуд - в головную аорту и далее выливается в полость тела. Через многочисленные отверстия гемолимфа попадает в полость перикардиального синуса, затем через остии, при расширении сердечной камеры, засасывается в сердце. Гемолимфа не имеет дыхательных пигментов и представляет желтоватую жидкость, содержащую фагоциты. Основная ее функция - снабжение органов питательными веществами и перенос продуктов обмена корганам выделения. Дыхательная функция гемолимфы незначительна, лишь у некоторых водных личинок насекомых (личинки комаров-звонцов) гемолимфа имеет гемоглобин, окрашена в ярко-красный цвет и отвечает за транспорт газов.
Органами выделения насекомых являются мальпигиевы сосуды и жировое тело. Мальпигиевы сосуды (в количестве до 150) имеют эктодермальное происхождение, впадают в просвет кишечника на границе между средним и задним отделами кишечника. Продукт выделения - кристаллы мочевой кислоты. Жировое тело насекомых помимо основной функции - накопления запасов питательных веществ, служит еще и «почкой накопления». В жировом теле имеются особые экскреторные клетки, которые постепенно насыщаются труднорастворимой мочевой кислотой.

рис. 7. Схема строения
кровеносной системы
черного таракана:
1 - сердце, 2 - аорта.
Центральная нервная система насекомых состоит из парных надглоточных ганглиев (головного мозга), подглоточных ганглиев и сегментарных ганглиев брюшной нервной цепочки. Головной мозг включает в себя три отдела: протоцеребрум, дейтоцеребрум и тритоцеребрум. Протоцеребрум иннервирует акрон и размещающиеся на нем глаза. На протоцеребруме развиваются грибовидные тела, к которым подходят нервы от органов зрения. Дейтоцеребрум иннервирует усики, тритоцеребрум - верхнюю губу.
В состав брюшной нервной цепочки входят 11-13 пар ганглиев: 3 грудных и 8-10 брюшных. У некоторых насекомых грудные и брюшные сегментарные ганглии сливаются в грудные и брюшные нервные узлы.
Периферическая нервная система представлена нервами, отходящими от центральной нервной системы, и органами чувств. Имеются нейросекреторные клетки, нейрогормоны которых регулируют деятельность эндокринных органов насекомых.
Чем сложнее поведение насекомых, тем сильнее развит у них головной мозг и грибовидные тела.
Органы чувств насекомых достигают высокой степени совершенства. Возможности их сенсорного аппарата часто превосходят таковые высших позвоночных животных и человека.
Органы зрения представлены простыми и сложными глазами (рис. 8). Сложные, или фасеточные глаза располагаются по бокам головы и состоят из омматидиев, число которых у различных видов насекомых варьирует от 8-9 (муравьи) до 28000 (стрекозы). У многих видов насекомых зрение цветное. Каждый омматидий воспринимает небольшую часть из поля зрения всего глаза, изображение складывается из множества мелких частичек изображения, такое зрение иногда называют «мозаичным». Роль простых глазков до конца не изучена, установлено, что они воспринимают поляризованный свет.

рис. 8.
А - фасеточный глаз (на разрезе видны омматидии), Б - схема
строения отдельного омматидия, В - схема строения простого
глазка: 1 - хрусталик, 2 - хрустальный конус, 3 - пигментные
клетки, 4 - зрительные (ретинальные) клетки,
5 - рабдом (зрительный стержень), 6 - фасетки (наружные
поверхности хрусталика), 7 - нервные волокна.
Многие насекомые способны издавать звуки и слышать их. Органы слуха и органы, издающие звуки, могут находиться на любом участке тела. Например, у кузнечиков, органы слуха (тимпанальные органы) располагаются на голенях передних ног, здесь имеются по две узкие продольные щели, ведущие к барабанной перепонке, связанной с рецепторными клетками. Органы, издающие звуки, находятся на передних крыльях, причем левое крыло соответствует «смычку», а правое - «скрипке».
Органы обоняния представлены совокупностью обонятельных сенсилл, расположенных, в основном, на антеннах. Антенны самцов развиты в большей степени, чем антенны самок. По запаху насекомые разыскивают пищу, места для откладки яиц, особей противоположного пола. Самки выделяют особые вещества - половые аттрактанты, привлекающие самцов. Самцы бабочек находят самок на расстоянии 3-9 км.
Вкусовые сенсиллы располагаются у жуков на челюстных и губных щупиках, у пчел, мух, бабочек - на ножках, у пчел, муравьев - на усиках.
Осязательные рецепторы, термо- и гигрорецепторы рассеяны по поверхности тела, но больше всего их на усиках и щупиках. Многие насекомые воспринимают магнитные поля и их изменение, где находятся органы, воспринимающие эти поля, пока неизвестно.
Насекомые - раздельнополые животные. У многих видов насекомых выражен половой диморфизм. В состав мужской половой системы входят: парные семенники и семяпроводы, непарный семяизвергательный канал, совокупительный орган и придаточные железы. В состав совокупительного органа входят кутикулярные элементы - гениталии. Придаточные железы выделяют секрет, разбавляющий сперму и формирующий оболочку сперматофора. В состав женской половой системы входят: парные яичник и яйцеводы, непарное влагалище, семяприемник, придаточные железы. Самки некоторых видов имеют яйцеклад. Гениталии самцов и самок имеют сложное строение и таксономическое значение.
Насекомые размножаются половым способом, для ряда видов известен партеногенез (тли).
Развитие насекомых делится на два периода - эмбриональное, включающее развитие зародыша в яйце, и постэмбриональное, которое начинается с момента выхода личинки из яйца и заканчивается смертью насекомого. Постэмбриональное развитие проходит с метаморфозом. По характеру метаморфоза эти членистоногие подразделяются на две группы: насекомые с неполным превращением (гемиметаболические) и насекомых с полным превращением (голометаболические).
У гемиметаболических насекомых личинка похожа на взрослое животное. Отличается от него недоразвитыми крыльями - гонадами, отсутствием вторичных половых признаков, меньшими размерами. Такие имагоподобные личинки называются нимфами. Личинка растет, линяет, после каждой линьки зачатки крыльев увеличиваются. После нескольких линек из нимфы старшего возраста выходит имаго.
У голометаболических насекомых личинка не похожа на имаго не только по строению, но и экологически, например, личинка майского жука живет в почве, имаго - на деревьях. Личинки после нескольких линек превращаются в куколок. Во время стадии куколки происходит разрушение личиночных органов и формирование организма взрослого насекомого.

рис. 9.
А - открытая (наездник), Б -
покрытая (бабочка),
В - скрытая (муха).
Личинки голометаболических насекомых не имеют сложных глаз и зачатков крыльев. Их ротовой аппарат грызущего типа, усики и конечности - короткие. По степени развития конечностей выделяют четыре типа личинок: протоподные, олигоподные, полиподные, аподные. У протоподных личинок имеются лишь зачатки грудных ног (пчелы). У олигоподных личинок - три пары нормальных ходильных ног (жуки, сетчатокрылые). Полиподные личинки кроме трех пар грудных ног имеют еще несколько пар ложных ножек на брюшке (бабочки, пилильщики). Брюшные ножки представляют собой выступы стенки тела, несут на подошве шипики и крючочки. Аподные личинки конечностей не имеют (двукрылые).
По способам движения личинки голометаболических насекомых подразделяются на камподеовидных, эруковидных, проволочниковых и червеобразных.
Камподеовидные личинки имеют длинное гибкое тело, бегательные ноги и чувствующие церки (жужелицы). Эруковидные личинки - мясистое слабоизогнутое телос конечностями или без (майские жуки, бронзовки, навозники). Проволочниковые - с жестким телом, круглым в поперечнике, с опорными церками (жуки-щелкуны, чернотелки). Червеобразные - по внешнему виду похожи на червей, безногие (двукрылые и многие другие).
Куколки бывают трех типов: свободные, покрытые, скрытые (рис. 9). У свободных куколок зачатки крыльев и конечностей хорошо видны, свободно отделены от тела, покровы - тонкие и мягкие (жуки). У покрытых куколок зачатки плотно прирастают к телу, покровы - сильно склеротизированные (бабочки). Скрытые куколки - свободные куколки, находящиеся внутри ложного кокона - пупария (мухи). Пупарий представляет собой несброшенную затвердевшую личиночную шкурку.
Способность видеть окружающий мир во всем спектре его цветов и оттенков - уникальный дар природы человеку. Мир красок, который способны воспринимать наши глаза, яркий и удивительный. Но человек не единственное живое существо на этой планете. Животные и насекомые также видят предметы, цвета, ночные очертания? Как видят мухи или пчелы нашу комнату, к примеру, или цветок?
Глаза насекомого
Современная наука с помощью специальных приборов сумела увидеть мир глазами разных животных. Это открытие стало сенсацией в свое время. Оказывается, многие братья наши меньшие, а особенно насекомые, видят совсем не ту картинку, которую наблюдаем мы. Видят ли мухи вообще? Да, но совсем не так, и получается, что мы и мухи, да и другие летающие и ползающие, живем вроде в одном мире, но совсем непохожем.
Все дело в У насекомых он не один или, вернее, не совсем один. Глаз насекомого - это собранные воедино тысячи или фасеток, или омматидий. Выглядят они как конусные линзы. Каждый такой омматидий видит разную, только ему доступную часть картинки. Как видят мухи? Изображение, которое они наблюдают, похоже на картинку, собранную из мозаики, или пазл.
Острота зрения насекомых зависит от количества омматидий. Самая зрячая - стрекоза, у нее омматидий - аж около 30 тысяч. Бабочки тоже зрячие - около 17 тысяч, для сравнения: у мухи - 4 тысяч, у пчелы - 5. Самый слабовидящий - муравей, его глаз вмещает всего 100 фасеток.
Круговая оборона
Еще одна способность насекомых, отличительная от человеческой, - возможность кругового обзора. Глаз-линза способен видеть все на 360 о. Среди млекопитающих самый большой угол зрения у зайца - 180 о. Поэтому он и прозван косым, а что делать, если врагов столько. Лев вот врагов не боится, и глаза у него рассматривают меньше 30 о горизонта. У маленьких насекомых природа компенсировала нехватку роста способностью видеть всех, кто к ним подкрадывается. Чем еще отличается зрительное восприятие насекомых, так это быстротой смены картинки. За время быстрого полета они успевают заметить все, что люди на такой скорости лицезреть не могут. Например, как видят мухи телевизор? Если бы наш глаз был таким, как у мухи или пчелы, крутить пленку нужно было бы в десять раз быстрее. Поймать муху сзади практически нереально, она видит взмах руки быстрее, чем он происходит. Человек кажется насекомым медлительной черепахой, а черепаха - вообще неподвижным камнем.

Цвета радуги
Практически все насекомые - дальтоники. Цвета они различают, но по-своему. Интересно, что глаза насекомых и даже некоторых млекопитающих не воспринимают красный цвет совсем или видят его как синий, фиолетовый. Для пчелы красные цветы выглядят черными. Растения, которым нужно опыление пчел, не цветут красным. Большинство ярких цветов алые, розовые, оранжевые, бордовые, но не красные. Те редкие, которые позволяют себе красный наряд, опыляются другим образом. Вот такая взаимосвязь в природе. Трудно представить, каким образом удалось ученым выяснить, как видят мухи расцветку комнаты, но оказывается, что любимым их цветом является желтый, а голубой и зеленый их раздражает. Вот так вот. Чтобы в кухне было мух меньше, просто нужно ее правильно покрасить.

Видят ли мухи в темноте?
Мухи, как большинство летающих насекомых, ночью спят. Да-да, им тоже нужен сон. Если муху постоянно сгонять и не давать ей спать на протяжении трех суток, она умирает. Мухи видят в темноте плохо. Это насекомые с круглыми глазами, но близорукие. Им не нужны глаза для поиска пищи.
В отличие от мух рабочие пчелы видят ночью хорошо, что позволяет им работать и в ночную смену тоже. Ночью цветы благоухают резче и соперников на нектар меньше.
Хорошо видят ночью но несомненным лидером по зрению в темноте признан американский таракан.
Форма предмета
Интересно восприятие формы предмета разными насекомыми. Специфика в том, что они могут совсем не воспринимать простые формы, которые не нужны для их жизнеспособности. Пчелы, бабочки не видят предметов простых форм, особенно неподвижных, зато их привлекает все, что имеет сложные формы цветов, особенно если они движутся, колышутся. Этим объясняется, в частности, и то, что пчелы и осы редко жалят стоящего неподвижно человека, а если и жалят, то в область губ, когда он разговаривает (движет губами). Мухи и некоторые другие насекомые человека не воспринимают, садятся они на него просто в поисках еды, которую ищут по запаху и видят датчиками на лапах.

Общие особенности зрения насекомых
- Красный цвет способны различить только бабочки - они и опыляют редкие цветы такой гаммы.
- Строение глаза у всех фасеточное, разница - в количестве омматидий.
- Трихромазия, или способность преображать цвета в три основных: фиолетовый, зеленый и ультрафиолетовый.
- Способность переламывать и отражать световые лучи и видеть картинку окружающей действительности целиком.
- Способность рассматривать картинки, которые меняются очень быстро.
- Насекомые умеют ориентироваться по солнечному свету, поэтому ночные бабочки слетаются к лампе.
- Бинокулярное зрение помогает хищникам в мире насекомых точно определять расстояния до своей жертвы.
Вернемся теперь к биологии. Человеческий глаз - отнюдь не единственный тип глаза. Хотя глаза почти всех позвоночных похожи на человеческие, однако у низших животных мы встречаем множество других типов глаз. Обсуждать их у нас нет времени. Но среди беспозвоночных (например, у насекомых) встречаются и высокоразвитые типы глаз; это сложные, или фасеточные, глаза. (У большинства насекомых, кроме больших: фасеточных глаз, есть еще простые глаза, или глазки.) Наиболее тщательно изучалось зрение пчелы. Изучать особенности зрения пчел легко, ибо их, как известно, привлекает мед и мы можем ставить опыты, смазывая медом, например, голубую или красную бумагу и наблюдая, какая из них привлечет пчелу. Этим методом были обнаружены очень интересные особенности зрения пчелы.
Прежде всего, пытаясь определить, насколько отчетливо пчела видит разницу между двумя кусочками «белой» бумаги, некоторые исследователи нашли, что она видит ее не очень хорошо, а другие, наоборот, что она делает это чертовски здорово. Даже если брались два почти в точности одинаковых кусочка бумаги, пчела все же различала их. Один кусок бумаги, например, отбеливался цинковыми белилами, а другой - свинцовыми, и, хотя оба они выглядели в точности одинаково, пчела различала их, ибо они по-разному отражают ультрафиолетовый свет. Таким образом, было обнаружено, что глаз пчелы чувствителен к более коротким длинам волн, чем глаз человека. Наши глаза видят в диапазоне от 7000 до 4000 Å, от красного до фиолетового, а пчелы могут видеть вплоть до 3000 Å, т. е. в ультрафиолетовой области! А это порождает целый ряд очень интересных эффектов. Прежде всего пчелы различают многие цветы, которые нам кажутся абсолютно одинаковыми. В этом нет ничего удивительного; ведь цветы цветут вовсе не для того, чтобы радовать наш взор. Они служат приманкой для пчел, своеобразным сигналом о том, что здесь есть мед. Всем известно, что есть очень много «белых» цветов. Цвет, который нам кажется белым, по-видимому, пчелы не видят, ибо выяснилось, что разные белые цветы не отражаютультрафиолетовые лучи столь полно, как это делаютистинно белые цветы. От белого предмета отражается не весь падающий на него свет, ультрафиолетовые лучи теряются, а это в точности то же, что для нас потеря голубого цвета, т. е. получение желтого цвета. Итак, все белые цветы кажутся пчелам цветными. Однако нам известно также, что пчелы не видят красного цвета. Выходит, можно считать, что красные цветы кажутся пчелам черными? Ничего подобного! Тщательное изучение красных цветов показывает, что, во-первых, даже наши глаза способны различать у огромного большинства красных цветов легкий синеватый оттенок, вызванный дополнительным отражением большинством из них синего цвета, который находится в видимой пчелами области. Кроме того, опыты также показывают, что цветы отличаются по своей способности отражать ультрафиолет от различных частей лепестка и т. д. Итак, если бы мы могли видеть цветы такими, как их видят пчелы, то нашли бы их еще прекраснее и разнообразнее!
Впрочем, было обнаружено, что имеются такие красные цветы, которые не отражают голубых или ультрафиолетовых лучей, поэтому они должны казаться пчелам черными! Это в какой-то степени объясняет недоумение тех людей, которых очень волнует этот вопрос: ведь черный цвет не кажется привлекательным, да и трудно его отличить от грязной густой тени. Так оно и получается на самом деле: пчелы не прилетают на эти цветы. Но зато как раз они нравятся крохотным колибри ; оказывается, эти птички отлично видят красный цвет!
Еще одна интересная сторона зрения пчелы. Взглянув на кусочек голубого неба и не видя самого солнца, пчела, по-видимому, может все-таки определить, где находится солнце. Для нас это не так-то просто. Посмотрите из окна на небо. Вы видите, что оно голубое. А в каком направлении находится сейчас солнце? Пчела может это определить, ибо она очень чувствительна к направлению поляризованного света, а отраженный от неба свет поляризован . До сих пор спорят о том, как это ей удается: то ли потому, что рассеяние света различно при разных обстоятельствах, то ли глаза пчелы непосредственно чувствительны к направлению поляризованного света. В самое недавнее время получены данные о непосредственной чувствительности глаза пчелы.
Говорят также, что пчела способна различать отдельные вспышки света с частотой 200 раз в 1 сев, тогда как мы различаем только 20 вспышек. В улье пчелы движутся очень быстро; они шевелят лапками, машут крылышками, но наши глаза с трудом успевают уследить за всеми этими движениями. Вот если бы мы могли различать более быстрые мелькания, тогда другое дело. Видимо, для пчелы очень важно, чтобы ее глаза имели столь быструю реакцию.
Теперь поговорим о том, какова, собственно, острота зрения у пчелы? Глаз пчелы сложный; состоит он из огромного числа особых глазков, называемых омматидиями, которые расположены на почти сферической поверхности по бокам головы насекомого.
На фиг. 36.7 показан омматидий. В его вершине находится прозрачная область, своего рода «хрусталик», но в действительности это больше напоминает фильтр, заставляющий свет идти вдоль узкого волокна, где, по-видимому, и происходит его поглощение. От другого его конца отходит нервное волокно. Центральное нервное волокно имеет по бокам шесть клеток, от которых по сути дела оно и отходит. Для наших целей этого описания вполне достаточно; главное то, что клетка имеет коническую форму, и множество таких клеток, примыкая друг к другу, образует поверхность глаза пчелы.
Посмотрим теперь, каково разрешение такого глаза. Проведем линию (фиг. 36.8), схематически представляющую омматидий, на поверхности глаза, который мы будем считать сферой радиусом r. Мы сейчас попытаемся вычислить ширину каждого омматидия, для чего напряжем немного нашу сообразительность и предположим, что природа столь же сообразительна, как и мы! Если омматидий очень велик, то разрешение не может быть большим. Иначе говоря, один омматидий получает информацию об одном направлении, соседний - о другом и т. д., а предметы, попавшие в промежуток, пчела не сможет видеть достаточно хорошо. Таким образом, неопределенность остроты зрения глаза, несомненно, связана с угловым размером конца омматидия относительно центра кривизны глаза. (На самом деле глаза расположены только на поверхности головы.) Но угол от одного омматидия до следующего равен, конечно, диаметру омматидия, деленному на радиус кривизны поверхности глаза:
Ф![]() иг.
36.7. Строение омматидия.
иг.
36.7. Строение омматидия.
Итак, можно сказать: «Чем меньше будет значение , тем больше окажется острота зрения.
Ф иг.
36.8. Схема распределения омматидиев на
поверхности глаза пчелы.
иг.
36.8. Схема распределения омматидиев на
поверхности глаза пчелы.
Но почему же тогда природа не дала пчеле очень-очень мелкие омматидии?» В ответ можно сказать следующее: мы уже достаточно хорошо знаем физику, чтобы понять, что при попытке пропустить свет через узкую щелку из-за дифракции невозможно достаточно хорошо видеть в данном направлении, ибо туда будет попадать свет от различных направлений, т. е. от всех направлений, находящихся внутри угла d , такого, что
(![]() 36.2)
36.2)
Теперь ясно, что, если взять б слишком маленьким, каждый омматидий вследствие дифракции будет видеть не только в одном направлении! Но если сделать значение слишком большим, то, хотя каждый будет смотреть в одном направлении, их окажется слишком мало, чтобы получить достаточно подробную картину. Таким образом, мы должны подобрать такое расстояние d, чтобы полный эффект этих двух механизмов был минимальным. Если мы сложим два выражения и найдем место, где сумма имеет минимум, то получим
( 36.3)
36.3)
что дает расстояние
(![]() 36.4)
36.4)
Вкнигах указывается диаметр, равный 30мк. Как видите, согласие оказывается довольно хорошим! Ясно, что именно этот механизм определяет размер глаза пчелы и он вполне доступен нашему пониманию. Подставив теперь полученное число в (36.1), легко определить, каково угловое разрешение глаза пчелы. Оно оказывается очень плохим по сравнению с глазом человека. Мы способны видеть вещи, кажущийся размер которых в тридцать раз меньше, чем видит пчела. Так что по сравнению с человеком изображение у пчелы получается довольно расплывчатое, несфокусированное.
Ф иг.
36.9. Оптимальный размер омматидия,
равный
m
.
иг.
36.9. Оптимальный размер омматидия,
равный
m
.
Тем не менее так оно есть, и на большее она просто не может рассчитывать. Естественно возникает вопрос: а почему бы пчеле не завести себе такой же глаз, как у нас, с хрусталиком и со всем прочим? Этому мешает несколько довольно интересных причин. Прежде всего пчела слишком мала; если бы у нее был глаз, похожий на наш, но соответственно уменьшенный, то размер зрачка оказался бы порядка 30 мк, а поэтому дифракция была бы так велика, что пчела все равно не видела бы лучше. Слишком маленький глаз - это не очень хорошо. Потом, если сделать глаз большим, как голова пчелы, то он занял бы всю голову. Ведь ценность сложного глаза в том и состоит, что он практически не занимает места - просто тоненький слой на поверхности головы пчелы. Так что, прежде чем давать советы пчеле, не забывайте, что у нее есть свои собственные проблемы!