Здесь мы рассмотрим лишь те вопросы, которые связаны со значением патологии клеточных мембран в развитии злокачественных новообразований. Хотя, по-видимому, нельзя считать злокачественную опухоль результатом только мембранной патологии, тем не менее в атипичных клетках отмечаются глубокие изменения мембран, которые могут таким образом становиться важным звеном патогенеза злокачественных образований.
1. Электронно-микроскопические исследования показали, что в клетках злокачественных опухолей наблюдаются эмбриональные особенности строения мембран, выражающиеся в слиянии между собой мембран внутриклеточных органелл и плазмолеммы. Мембранная поверхность клетки таким образом резко увеличивается, что в конечном итоге приводит к существенным изменениям транспортных процессов, питания клетки, восприятия мембранами информации и т.д.
2. В одной мембране нормальных клеток может существовать более сотни неодинаковых по структуре липидных веществ, причем соотношения между ними специфичны для определенного типа мембран. Каждая мембрана имеет присущий только ей липидный состав. Во многих опухолевых клетках эти различия в липидном компоненте нивелируются или даже полностью отсутствуют. Наблюдается монотонность липидной структуры мембран, причем она выражена тем сильнее, чем быстрее растет опухоль и чем она злокачественнее.
3. Американский исследователь Коумен установил факт ослабления силы сцепления между опухолевыми клетками: она оказалась в десять раз меньшей, чем для нормальных клеток той же ткани.
Электронно-микроскопический анализ показал, что у опухолевых клеток имеется меньше участков сцепления друг с другом, чем у нормальных. Предполагают, что сцепление клеток происходит благодаря контакту фиксированного на мембранах кальция с отрицательно заряженными молекулярными группировками на мембране другой клетки. В качестве доказательства роли кальция в этом сцеплении можно привести наблюдения, показавшие, что обработка эмбрионов амфибий ЭДТА (вещество, связывающее ионы кальция) приводила к распаду ткани на отдельные клетки. Когда же к этим клеткам добавляли ионы кальция, они вновь объединялись между собой и продолжали эмбриональное развитие.
Поскольку клетки перестают сцепливаться между собой при их обработке трипсином, можно предположить, что они склеиваются и белковыми компонентами. Швейцарский исследователь Бюргер на клетках губок показал, что важную роль в этом склеивании играют ионы кальция и так называемый фактор агрегации, представляющий собой сложный комплекс белков и сахаристых веществ. Альберт Сент-Дьерди отводит важную роль в этом сцеплении глиоксалевой кислоте, поскольку сцепление клеток ослабевает при обработке их ферментом глиоксалазой, а добавление к опухолевым клеткам метилглиоксаля усиливает их контакт между собой. Предполагают также, что рецепторы контактов у опухолевых клеток более подвижны, так как они «плавают» в более жидких липидах, поэтому они могут сгруппировываться, и число межклеточных контактов таким образом уменьшается.
4. Процесс сцепления нормальных клеток между собой приводит еще к одному феномену - контактному торможению. Две нормальные клетки, будучи помещенными в каплю жидкости и передвигаясь в ней, при контакте и сцеплении между собой перестают двигаться; кроме того в них начинает тормозиться синтез ДНК, в результате чего они перестают делиться и расти. Чем больше клеток сцеплено друг с другом, тем сильнее тормозится их рост и деление. У опухолевых клеток эти явления не наблюдаются. В чем причина отсутствия у опухолевых клеток контактного торможения? В потере способности к передаче сигнала, останавливающего рост и деление, или в отсутствии способности воспринимать такой сигнал? Опыты американского исследователя Стоукера проливают некоторый свет на эту проблему. Он показал, что при смешении нормальных фибробластов в культуре тканей с трансформированными в последних наблюдалось торможение деления и роста. Таким образом, дело, по-видимому, заключается в потере опухолевыми клетками способности к генерации соответствующего сигнала. Отсюда следует, что малое количество опухолевых клеток еще может быть нормализовано обычным клеточным окружением, а большое - нет. Другими словами, для того чтобы развилась опухоль, количество опухолевых клеток должно достигнуть определенной критической величины.
Проницаемость мембран опухолевых клеток по сравнению с нормальными является повышенной, по-видимому, из-за сокращения количества внеклеточных контактов и увеличения свободной мембранной поверхности. Вследствие этого в бластомных клетках усиливаются транспортные процессы и происходит более быстрое «изнашивание» гиперфункционирующих мембранных насосов.
Патология клетки
Клетка – элементарная живая система, обладающая способностью к обмену с окружающей средой. Строение клеток организма человека обеспе-чивает выполнение ими специализированной функции и «сохранение себя», т. е. поддержание клеточного пула. Органоиды клетки, обладая определенными морфологическими особенностями, обеспечивают основные проявления жизнедеятельности клетки. С ними связаны дыхание и энергетические запасы (митохондрии), синтез белков (рибосомы, гранулярная цитоплазматическая сеть), накопление и транспорт липидов и гликогена, детоксикационная функция (гладкая цитоплазматическая сеть), синтез продуктов и их секреция (пластинчатый комплекс), внутриклеточное пищеварение и защитная функция (лизосомы). Деятельность ультраструктур клетки строго координирована, причем координация в выработке специфического продукта клеткой подчинена закону «внутриклеточного конвейера». По принципу ауторегуляции он осуществляет взаимосвязь между структурными компонентами клетки и протекающими в ней процессами обмена.
Функции органоидов не строго детерминированы, так как они могут участвовать в различных внутриклеточных процессах. Более специализированы метаплазматические образования клетки, выполняющие частные функции: тонофибриллы, выполняющие опорную функцию клетки; миофибриллы, осуществляющие сокращение клетки и способствующие ее движению; микроворсинки, щеточная каемка, участвующие в процессах всасывания; десмосомы, обеспечивающие клеточные контакты, и т. д. Однако ни одна функция клетки не является результатом деятельности одного органоида или одного метаплазматического образования. Каждое функциональное проявление клетки – это результат совместной работы всех взаимосвязанных компонентов. Понятно поэтому, что структурные изменения клетки, отражающие нарушения ее функции, не могут быть поняты без учета возможных изменений каждой из ее двух основных частей – ядра и цитоплазмы, ее органелл, метаплазматических образований и включений. От нарушений элементарных структур клетки и их функций к патологии клетки как элементарной саморегулирующейся живой системе и к патологии клеточных коопераций, объединенных конечной функцией,- таков путь познания патологии клетки – структурной основы патологии человека.
Поэтому патология клетки – понятие неоднозначное. Во-первых, это патология специализированных ультраструктур клетки, она представлена не только достаточно стереотипными изменениями той или иной ультраструктуры в ответ на различные воздействия, но и настолько специфичными изменениями ультраструктур, что можно говорить о хромосомных болезнях и «болезнях» рецепторов, лизосомных, митохондриальных, пероксисомных и других «болезнях» клетки. Во-вторых, патология клетки – это изменения ее компонентов и ультраструктур в причинно-следственных связях. При этом речь идет о выявлении общих закономерностей повреждения клетки и ее реакции на повреждение. Сюда могут быть отнесены: рецепция патогенной информации клеткой и реакция на повреждение, нарушения проницаемости клеточных мембран и циркуляции внутриклеточной жидкости; нарушения метаболизма клетки, смерть клетки (некроз), клеточная дисплазия и метаплазия, гипертрофия и атрофия, патология движения клетки, ее ядра и генетического аппарата и др.
Патология клеточного ядра
Морфологически она проявляется в изменении структуры, размеров, формы и количества ядер и ядрышек, в появлении разнообразных ядерных включений и изменений ядерной оболочки. Особую форму ядерной патологии представляет патология митоза; с патологией хромосом ядра связано развитие хромосомных синдромов и хромосомных болезней.
Структура и размеры ядер
Структура и размеры ядра (речь идет об интерфазном, интермитозном, ядре) зависят в первую очередь от плоидности, в частности от содержания в ядре ДНК, и от функционального состояния ядра. Тетраплоидные ядра имеют диаметр больше, чем диплоидные, октоплоидные – больше, чем тетраплоидные.
Большая часть клеток содержит диплоидные ядра. В пролиферирующих клетках в период синтеза ДНК (S-фаза) содержание ДНК в ядре удваивается, в постмитотический период, напротив, снижается. Если после синтеза ДНК в диплоидной клетке не происходит нормального митоза, то появляются тетраплоидные ядра. Возникает полиплоидия – кратное увеличение числа наборов хромосом в ядрах клеток, или состояние плоидности от тетраплоидии и выше.
Полиплоидные клетки выявляют различными способами: по размеру ядра, по увеличенному количеству ДНК в интерфазном ядре или по увеличению числа хромосом в митотической клетке. Они встречаются в нормально функционирующих тканях человека. Увеличение числа полиплоидных ядер во многих органах отмечается в старости. Особенно ярко полиплоидия представлена при репаративной регенерации (печень), компенсаторной (регенерационной) гипертрофии (миокард), при опухолевом росте.
Другой вид изменений структуры и размеров ядра клетки встречается при анеуплоидии, под которой понимают изменения в виде неполного набора хромо-сом. Анеуплоидия связана с хромосомными мутациями. Ее проявления (гипертетраплоидные, псевдоплоидные, «приблизительно» диплоидные или триплоидные ядра) часто обнаруживаются в злокачественных опухолях.
Размеры ядер и ядерных структур независимо от плоидии определяются в значительной мере функциональным состоянием клетки. В связи с этим следует помнить, что процессы, постоянно совершающиеся в интерфазном ядре, разнонаправленны: во-первых, это репликация генетического материала в S-нериоде («полуконсервативный» синтез ДНК); во-вторых, образование РНК в процессе транскрипции, транспортировка РНК из ядра в цитоплазму через ядерные поры для осуществления специфической функции клетки и для репликации ДНК.
Функциональное состояние ядра находит отражение в характере и распределении его хроматина. В наружных отделах диплоидных ядер нормальных тканей находят конденсированный (компактный) хроматин – гетерохроматин, в остальных ее отделах – неконденсированный (рыхлый) хроматин – эухроматин. Гетеро- и эухроматин отражают различные состояния активности ядра; первый из них считается «малоактивным» или «неактивным», второй – «достаточно активным». Поскольку ядро может переходить из состояния относительно функционального покоя в состояние высокой функциональной активности и обратно, морфологическая картина распределения хроматина, представленная гетеро- и эухроматином, не может считаться статичной. Возможна «гетерохроматинизация» или «эухроматинизация» ядер, механизмы которой изучены недостаточно. Неоднозначна и трактовка характера и распределения хроматина в ядре.
Например, маргинация хроматина, т. е. расположение его под ядерной оболочкой, трактуется и как признак активности ядра, и как проявление его повреждения. Однако конденсация эухроматиновых структур (гиперхроматоз стенки ядра), отражающая инактивацию активных участков транскрипции, рассматривается как патологическое явление, как предвестник гибели клетки. К патологическим изменениям ядра относят также его дисфункциональное (токсическое) набухание, встречающееся при различных повреждениях клетки. При этом происходит изменение коллоидно-осмотического состояния ядра и цитоплазмы вследствие торможения транспорта веществ через оболочку клетки.
Форма ядер и их количество
Изменения формы ядра – существенный диагностический признак: деформация ядер цитоплазматическими включениями при дистрофических процессах, полиморфизм ядер при воспалении (гранулематоз) и опухолевом росте (клеточный атипизм).
Форма ядра может меняться также в связи с образованием множественных выпячиваний ядра в цитоплазму (рис. 3), которое обусловлено увеличением ядерной поверхности и свидетельствует о синтетической активности ядра в отношении нуклеиновых кислот и белка.
Изменения количества ядер в клетке могут быть представлены многоядерностью, появлением «спутника ядра» и безъядерностью. Многоядерность возможна при слиянии клеток. Таковы, например, гигантские многоядерные клетки инородных тел и Пирогова – Лангханса, образующиеся при слиянии эпителиоидных клеток (см. рис. 72). Но возможно образование многоядерных клеток и при нарушениях митоза – деление ядра без последующего деления цитоплазмы, что наблюдается после облучения или введения цитостатиков, а также при злокачественном росте.
«Спутниками ядра», кариомерами (маленькими ядрами), называют мелкие подобные ядру образования с соответствующей структурой и собственной оболочкой, которые расположены в цитоплазме около неизмененного ядра. Причиной их образования считают хромосомные мутации. Таковы кариомеры в клетках злокачественной опухоли при наличии большого числа фигур патологических митозов.
Безъядерность в отношении функциональной оценки клетки неоднозначна. Известны безъядерные клеточные структуры, которые являются вполне жизнеспособными (эритроциты, тромбоциты). При патологических состояниях можно наблюдать жизнеспособность частей цитоплазмы, отделенных от клетки. Но безъядерность может свидетельствовать и о гибели ядра, которая проявляется кариопикнозом, кариорексисом и кариолизисом (см. Некроз).
Структура и размеры ядрышек
Изменения ядрышек имеют существенное значение в морфофункциональной оценке состояния клетки, так как с ядрышками связаны процессы транскрипции и трансформации рибосомальной РНК (р-РНК). Размеры и структура ядрышек в большинстве случаев коррелируют с объемом клеточного белкового синтеза, выявляемого биохимическими методами. Размеры ядрышек зависят также от функции и типа клеток.
Увеличение размеров и количества ядрышек свидетельствует о повышении их функциональной активности. Вновь образованная в ядрышке рибосомальная РНК транспортируется в цитоплазму и, вероятно, через поры внутренней ядерной мембраны. Интенсивный синтез белка в таких случаях подтверждается увеличением количества рибосом эндоплазматической сети.
Гипергранулированные ядрышки с преобладанием гранул над фибриллярной субстанцией могут отражать различное функциональное состояние как ядрышек, так и клетки. Наличие таких ядрышек с хорошо выраженной лакунарной системой и резкой базофилией цитоплазмы свидетельствует как о повышенном синтезе р-РНК, так и о трансмиссии. Такие «гиперфункциональные ядрышки» встречаются в молодых плазматических клетках, активных фибробластах, гепатоцитах, во многих опухолевых клетках. Те же гипергранулированные ядрышки со слабовыраженной базофилией цитоплазмы могут отражать нарушение трансмиссии (транспортировки гранул) при продолжающемся синтезе р-РНК. Они обнаруживаются в опухолевых клетках, отличающихся большим ядром и незначительной цитоплазматической базофилией.
Разрыхление (диссоциация) ядрышек, отражающее их гипогрануляцию, может быть следствием «извержения» р-РНК в цитоплазму или торможения ядрышковой транскрипции. Дезорганизация (сегрегация) ядрышек отражает, как правило, полное и быстрое прекращение ядрышковой транскрипции: ядро уменьшается в размерах, наблюдается выраженная конденсация ядрышкового хроматина, происходит разделение гранул и протеиновых нитей. Эти изменения встречаются при энергетическом дефиците клетки.
Ядерные включения
Ядерные включения делят на три группы:
1. ядерные цитоплазматические
2. истинные ядерные
3. ядерные вирусобусловленные.
Ядерными цитоплазматическими включениями называют отграниченные оболочкой части цитоплазмы в ядре. Они могут содержать все составные части клетки (органеллы, пигмент, гликоген, капли жира и т. д.). Их появление в большинстве случаев связано с нарушением митотического деления.
Истинными ядерными включениями считают те, которые расположены внутри ядра (кариоплазмы) и соответствуют веществам, встречающимся в цитоплазме [белок, гликоген, липиды и т. д.]. В большинстве случаев эти вещества проникают из цитоплазмы в ядро через неповрежденные или поврежденные поры ядерной оболочки или через разрушенную ядерную оболочку. Возможно также проникновение этих веществ в ядро при митозе. Таковы, например, включения гликогена в ядрах печени при сахарном диабете («ядерный гликоген», «дырчатые, пустые, ядра»).
Вирусобусловленные ядерные включения (так называемые тельца ядерных включений) неоднозначны. Во-первых, это ядерные включения в кариоплазме кристаллической решетки вируса, во-вторых, включения белковых частиц, возникающих при внутриядерном размножении вируса; в-третьих, ядерные включения как проявление реакции на поражение вирусом цитоплазмы («реактивные включения»).
Ядерная оболочка
Ядерная оболочка выполняет ряд функций, нарушения которых могут служить основой для развития патологии клетки.
О роли ядерной оболочки в поддержании формы и размера ядра свидетельствует образование внутриядерных трубчатых систем, отходящих от внутренней ядерной мембраны, включений в перинуклеарной зоне [гипертрофия миокарда, легочный фиброз, системный васкулит, саркоидоз, опухоли печени, дерматомиозит].
О ядерной оболочке как месте прикрепления ДНК для облегчения репликации и транскрипции свидетельствует тот факт, что в ядерной оболочке имеются структуры, модулированные хроматином и в свою очередь ответственные за ориентацию и структуру хроматина. Показано, что функциональная активность ДНК связана с ее распределением при делении клетки и со степенью конденсации в интерфазе, причем повреждение оболочки может вызывать изменения таких участков распределения и быть причиной патологических изменений клетки.
В пользу функции ядерной оболочки как физического барьера и модулятора нуклеоцитоплазматического обмена говорит установленная корреляция между изменениями структуры ядерной оболочки, модулем ее пор и выходом РНК в цитоплазму. Контроль ядерной оболочкой транспорта РНК в цитоплазму может оказывать существенное влияние на гомеостаз клетки при патологических состояниях. Участие ядерной оболочки в синтезе мембран не имеет достоверных доказательств, хотя и считают, что эта роль возможна, так как мембраны ядерной оболочки непосредственно переходят в эндоплазматическую сеть цитоплазмы. О возможном влиянии ферментов ядерной оболочки на функцию ядра свидетельствует наличие в ядерной оболочке различных ферментов детоксикации, а. также веществ, обеспечивающих «гормон,альное управление» (аденилатциклаза, рецепторы инсулина и др.).
Патология митоза
В жизненном цикле клетки митоз занимает особое место. С его помощью осуществляется репродукция клеток, а значит, и передача их наследственных свойств. Подготовка клеток к митозу складывается из ряда последовательных процессов: репродукции ДНК, удвоения массы клетки, синтеза белковых компонентов хромосом и митотического аппарата, удвоения клеточного центра, накопления энергии для цитотомии. В процессе митотического деления, как известно, различают 4 основные фазы: профазу, метафазу, анафазу и телофазу.
При патологии митоза может страдать любая из этих фаз. Руководствуясь этим, создана классификация патологии митоза [Алов И. А., 1972], согласно которой выделяются следующие типы патологии митоза:
I. Повреждение хромосом:
1. задержка клеток в профазе;
2. нарушение спирализации и деспирализации хромосом;
3. фрагментация хромосом;
4. образование мостов между хромосомами в анафазе;
5. раннее разъединение сестринских хроматид;
6. повреждение кинетохора.
II. Повреждение митотического аппарата:
1. задержка развития митоза в метафазе;
2. рассредоточение хромосом в метафазе;
3. трехгрупповая метафаза;
4. полая метафаза;
5. многополюсные митозы;
6. асимметричные митозы;
7. моноцентрические митозы;
8. К-митозы.
III. Нарушение цитотомии:
1. преждевременная цитотомия;
2. задержка цитотомии;
3. отсутствие цитотомии.
Патологию митоза могут вызвать различные воздействия на клетку: ультрафиолетовое и ионизирующее излучение, высокая температура, химические вещества, в том числе канцерогены и митотические яды и др. Велико количество патологических митозов при малигнизации тканей.
Хромосомные аберрации и хромосомные болезни
Хромосомные аберрации.
Под хромосомными аберрациями понимают изменения структуры хромосом, вызванные их разрывами, с последующим перераспределением, утратой или удвоением генетического материала. Они отражают различные виды аномалий хромосом. У человека среди наиболее часто встречающихся хромосомных аберраций, проявляющихся развитием глубокой патологии, выделяют аномалии, касающиеся числа и структуры хромосом. Нарушения числа хромосом могут быть выражены отсутствием одной из пары гомологичных хромосом (моносомия) или появлением добавочной, третьей, хромосомы (трисомия). Общее количество хромосом в кариотипе в этих случаях отличается от модального числа и равняется 45 или 47. Полиплоидия и анеуплоидия имеют меньшее значение для развития хромосомных синдромов. К нарушениям структуры хромосом при общем нормальном их числе в кариотипе относят различные типы их «поломки»: транслокадию (обмен сегментами между двумя негомологичными хромосомами), делецию (выпадение части хромосомы), фрагментацию, кольцевые хромосомы и т. д.
Хромосомные аберрации, нарушая баланс наследственных факторов, являются причиной многообразных отклонений в строении и жизнедеятельности организма, проявляющихся в так называемых хромосомных болезнях.
Хромосомные болезни.
Их делят на связанные с аномалиями соматических хромосом (аутосом) и с аномалиями половых хромосом (телец Барра). При этом учитывают характер хромосомной аномалии – нарушение числа отдельных хромосом, числа хромосомного набора или структуры хромосом. Эти критерии позволяют выделять полные или мозаичные клинические формы хромосомных болезней.
Хромосомные болезни, обусловленные нарушениями числа отдельных хромосом (трисомиями и моносомиями), могут касаться как аутосом, так и половых хромосом.
Моносомии аутосом (любые хромосомы, кроме Х- и Y-хромосом) несовместимы с жизнью. Трисомии аутосом достаточно распространены в патологии человека. Наиболее часто они представлены синдромами Патау (13-я пара хромосом) и Эдвардса (18-я пара), а также болезнью Дауна (21-я пара). Хромосомные синдромы при трисомиях других пар аутосом встречаются значительно реже. Моносомия половой Х-хромосомы (генотип ХО) лежит в основе синдрома Шерешевского-Тернера, трисомия половых хромосом (генотип XXY) – в основе синдрома Клейнфелтера. Нарушения числа хромосом в виде тетра- или триплоидии могут быть представлены как полными, так и мозаичными формами хромосомных болезней.
Нарушения структуры хромосом дают самую большую группу хромосомных синдромов (более 700 типов), которые, однако, могут быть связаны не только с хромосомными аномалиями, но и с другими этиологическими факторами.
Для всех форм хромосомных болезней характерна множественность проявлений в виде врожденных пороков развития, причем их формирование начинается на стадии гистогенеза и продолжается в органогенезе, что объясняет сходство клинических проявлений при различных формах хромосомных болезней.
Патология цитоплазмы
Изменения мембран и патология клетки
Клеточные мембраны, как известно, состоят из бислоя фосфолипидов, по обе стороны которого располагаются разнообразные мембранные белки. На внешней поверхности мембраны белковые молекулы несут полисахаридные компоненты (гликокаликс), которые содержат многочисленные поверхностные клеточные антигены. Они играют важную роль в формировании клеточных стыков.
Изменения клеточных мембран.
Среди них различают следующие [Авцын А. П., Шахламов В. А., 1979]: чрезмерное везикулообразование («минус-мембрана»); увеличение поверхности плазмолеммы клеток мембранами микропиноцитозных пузырьков («плюс-мембрана»); усиленный микроклазматоз и клазматоз («минус-мембрана»); образование цитоплазматических отростков из плазмолеммы клетки; образование пузырей на поверхности клетки; утолщение слоев мембраны; образование микропор; образование миелиноподобных структур из плазмолеммы и мембран органелл; слияние разнородных клеточных мембран; локальные разрушения мембран – «бреши» в плазмолемме; «штопка» локально разрушенной плазмолеммы мембранами микропиноцитозных везикул.
К патологии мембран клетки могут вести нарушения мембранного транспорта, изменения проницаемости мембран, изменения коммуникации клеток и их «узнавания», изменения подвижности мембран и формы клеток, нарушения синтеза и обмена мембран.
Нарушения мембранного транспорта.
Процесс мембранного транспорта предполагает перенос ионов и других субстратов против градиента концентрации. Транспорт может быть активным, тогда он требует АТФ и «подвижности» транспортных белков в мембране, или пассивным посредством различных диффузионных и обменных процессов. Активный транспорт – это также функция эпителиальных барьеров. Нарушения мембранного транспорта, ведущие к патологии клетки, хорошо прослежены при ишемии, которая приводит к первичным изменениям митохондрий. В митохондриях резко падает эффективность окислительного фосфорилирования, они набухают, вначале увеличивается проницаемость их внутренней мембраны, в дальнейшем повреждение становится тотальным и необратимым.
Ишемическое повреждение митохондрий приводит к полому натрий-калиевого АТФ-насоса, постепенному накапливанию в клетке натрия и потере ею калия. Нарушение натрий-калиевого обмена ведет к вытеснению кальция из митохондрий. В результате в цитоплазме повышается уровень ионизированного кальция и увеличивается связывание его с кальмодулином. С повышением содержания кальций-кальмодулиновых комплексов связан ряд изменений клетки: расхождение клеточных стыков, поглощение кальция митохондриями, изменение микротрубочек и микрофиламентов, активация фосфолипаз. Эндоплазматическая сеть накапливает воду и ионы, следствием чего является расширение ее канальцев и цистерн, развитие гидропической дистрофии. Усиление гликолиза сопровождается истощением гликогена, накоплением лактата и снижением клеточного рН. С этими изменениями связано нарушение структуры хроматина и уменьшение синтеза РНК. Необратимые ишемические повреждения клетки связаны с гидролизом мембран, особенно мембранных липидов, под действием фосфолипаз. Возникают и нарушения лизосомальных мембран с высвобождением гидролаз.
Изменения проницаемости мембран.
Контроль мембранной проницаемости предполагает поддержание структуры как фосфолипидного бислоя мембраны с необходимым обменом и ресинтезом, так и соответствующих белковых каналов. Важная роль в осуществлении этого контроля принадлежит гликокаликсу и взаимодействию мембранных белков с цитоскелетом, а также гормонам, взаимодействующим с мембранными рецепторами. Изменения проницаемости могут быть тяжелыми (необратимыми) или поверхностными. Наиболее изученной моделью изменения мембранной проницаемости является повреждение тяжелыми металлами (ртуть, уран). Тяжелые металлы, взаимодействуя с сульфгидрильными группами мембранных белков, изменяют их конформацию и резко увеличивают проницаемость мембраны для натрия, калия, хлора, кальция и магния, что приводит к быстрому набуханию клеток, распаду их цитоскелета. Подобные изменения мембран отмечаются при повреждении их комплементом («болезни гиперчувствительности»). В мембранах образуются бреши, что снижает их сопротивление и резко увеличивает проницаемость.
Изменения коммуникации клеток и их «узнавания». Коммуникабельность клеток и опознавание «своих» и «чужих» – необходимое свойство клеточного кооперирования. Клеточное «общение» и «узнавание» подразумевают прежде всего различия во внешних поверхностях плазматической мембраны н мембран внутриклеточных органелл. Особый интерес в этом отношении представляет гликокаликс мембраны с поверхностными антигенами – маркерами определенного типа клеток.
Изменения клеточного «общения» и «узнавания» встречаются при тех патологических процессах (воспаление, регенерация, опухолевый рост), при которых поверхностные антигены могут изменяться, причем различия могут касаться как типа антигена, так и его «доступности» со стороцы внеклеточного пространства. Показано, что при исчезновении характерных для данного типа клеток антигенов могут появляться «эмбриональные» и аномальные (например, карциноэмбриональный) антигены; изменения гликолипидов мембраны делают ее более доступной воздействию антител.
Коммуникабельность клеток определяется также состоянием клеточных стыков, которые могут повреждаться при различных патологических процессах и болезнях. В раковых клетках, например, найдена корреляция между изменениями клеточных стыков и нарушением межклеточных связей; в опухолях обнаружены аномальные клеточные соединения.
Изменения подвижности мембран и формы клеток. Различают два типа изменений, связанных с нарушением подвижности мембран: выпячивание мембраны наружу – экзотропия и внутрь цитоплазмы – эзотропия. При экзотропии мембрана, выпячивающаяся во внеклеточное пространство, образует окруженную мембраной цитоплазматическую структуру. При эзотропии появляется окруженная мембраной полость. Изменения формы клеток связаны не только с экзо- и эзотропией, но и с упрощением клеточной поверхности (потеря малых отростков подоцитов при нефротическом синдроме).
Нарушения синтеза и обмена мембран. Возможно усиление синтеза мембран (при воздействии ряда химических веществ на клетку) или его ослабление (снижение синтеза мембран щеточной каемки энтероцитов при угнетении мембранных ферментов). В равной мере возможно усиление обмена мембран (при стимуляции аутофагоцитоза) или его ослабление (при лизосомных болезнях).
Эндоплазматическая сеть
Изменения гранулярной эндоплазматической сети и рибосом
Функции гранулярной эндоплазматической сети и рибосом сопряжены достаточно жестко, поэтому морфологические проявления их нарушений касаются, как правило, обеих органелл.
Изменения гранулярной эндоплазматической сети и рибосом могут быть представлены гиперплазией и атрофией, упрощением структуры, дезагрегацией (диссоциацией) рибосом и полисом, образованием аномальных рибосомально-пластинчатых комплексов.
Гиперплазия гранулярной эндоплазматической сети и рибосом, т. е. увеличение их количества, светооптически проявляется повышенной базофилией цитоплазмы, которая отражает объемную плотность рибосом и является показателем интенсивности белкового синтеза в клетке. Электронно-микроскопически в таких случаях можно судить о сопряжении синтеза и экскреции белка или отсутствии такого сопряжения. В интенсивно секретирующих и экскретирующих белок клетках (например, в активных фибробластах) цистерны гранулярной эндоплазматической сети расширены и содержат мало электронно-плотного материала: отмечается гиперплазия как связанных с мембранами, так и свободных рибосом, образующих полисомы; пластинчатый комплекс (комплекс Гольджи), участвующий в экскреции синтезируемого белка, хорошо развит. В интенсивно секретирующих белок клетках с нарушенной его экскрецией в гиперплазированных расширенных цистернах эндоплазматической сети с обилием рибосом и полисом накапливается хлопьевидный электронно-плотный материал, иногда происходит его кристаллизация; комплекс Гольджи в таких случаях развит плохо.
Как отдельные клетки, так и целые многоклеточные организмы могут подвергаться различным воздействиям, которые приводят к их структурно-функциональным изменениям, к нарушениям их жизненных функций – патологии.
Патологические изменения одноклеточных организмов, приводящие к временным нарушениям их отдельных функций или приводящие к стойким нарушениям, кончающимся гибелью этой клетки - организма, есть результат поражения отдельных внутриклеточных структур.
В многоклеточных организмах также по ряду разнообразных причин происходят изменения или повреждения группы клеток, которые могут приводить к развитию целого набора дополнительных функциональных нарушений уже вторичного характера, связанных с изменением других клеток, так развивается патологические изменения целого организма, развивается болезнь как системное нарушение целого ряда клеток и тканей.
Изучение различных типов клеточного поражения, процессов их развития, способности клеток к репаративным процессам имеет большое общебиологическое значение, раскрывая пути взаимосвязи и регуляции между отдельными клеточными компонентами и прикладное значение, так как прямо связано с задачами медицины.
Современная биология рассматривает клетку как единую комплексную, интегрированную систему, где отдельные функции взаимосвязаны и сбалансированы друг с другом. Поэтому нарушение и выпадение отдельных этапов клеточного метаболизма должны приводить или к активации запасных обходных путей или к развертыванию событий уже патологического характера. У многоклеточных организмов патология и гибель ряда клеток используются в здоровом организме в целях прогрессивных нормальных процессов. В этом случае происходит запрограммированное выключение тех или иных клеточных функций, приводящих к смерти клетки.
Более подробно изучено действие на различные клетки внешних повреждающих факторов, физических и химических, таких как температура, лучистая энергия, давление, действие неспецифических альтерирующих химических веществ и влияние ингибиторов отдельных звеньев клеточного метаболизма и антибиотиков.
Разнообразные факторы при обратимом повреждении клетки отвечают ограниченным числом неспецифических изменений. Наблюдения привели к выводу, что эти морфофункциональные показатели повреждения возникают стереотипно вне зависимости от природы клеток или от типа повреждающего фактора. Неспецифический характер ответных реакций клетки на различные повреждающие факторы может свидетельствовать о наличии, каких – то общих процессов, вызывающих развитие сходных ответов клеток. При этом в клетках всегда значительно падает окислительное фосфорилирование, возрастание гликолитических процессов, активация протеолиза.
Характерной общеклеточной реакцией на повреждение является изменение способности клетки связывать различные красители. В морфологическом отношении начинают появляться структурно–патологические изменения: распад вакуолярной системы, активация лизосом, изменение структуры митохондрий и ядра. Совокупность неспецифических обратимых изменений цитоплазмы, возникающих под воздействием различных агентов, была обозначена термином «паранекроз».
К патологическим процессам на клеточном уровне относятся не только явления, связанные деструкцией клеток. Другой уровень клеточной патологии – изменение регуляторных процессов. Это могут быть нарушения регуляции обменных процессов, приводящие к отложению различных веществ, нарушения дифференцировки (например, опухолевый рост).
Некоторые термины:
пролиферация (proliferatio; лат. proles потомство + fero несу, приношу) - увеличение числа клеток какой-либо ткани вследствие их размножения;
пролиферативный пул - отношение количества размножающихся клеток ко всей массе данной клеточной популяции;
репродукция (ре- + лат. productio производство) - 1) в биологии = Размножение; 2) в психологии = Воспроизведение;
кариотип (карио- греч. karyon ядро, орех + греч. typos форма, образец) -- совокупность морфологических особенностей хромосомного набора соматической клетки организма данного биологического вида;
ген (-ы) (греч. genos род, рождение, происхождение) -- структурная и функциональная единица наследственности, контролирующая образование какого-либо признака, представляющая собой отрезок молекулы дезоксирибонуклеиновой кислоты (у некоторых вирусов -- рибонуклеиновой кислоты);
генотип (ген + греч. typos отпечаток, образец, тип; син.: идиотип, конституция генетическая) -- совокупность всех генов, присущих данной особи;
геном (англ. genome, от греч. genos род, происхождение) -- совокупность хромосомных наследственных факторов, передаваемых от родительской особи к дочерней, представляющая собой у эукариотов, в том числе у человека, гаплоидный набор хромосом;
Контрольные вопросы:
1. Механизм и роль амитоза
2. Значение митоза для клетки
3. Этапы митоза
4. Роль центросомы в делении клетки
5. Этапы мейоза
6. Первое деление мейоза
7. Второе деление мейоза
8. Роль кроссинговера в модифицировании индивидуальной наследственной информации
9. Отличия митоза и мейоза, их биологическое значение
10. Клеточный цикл, его фазы и регуляция
1. Альбертс Б., Брей Д., Льюис Д. И др. Молекулярная биология клетки: В 3-х тт. -М., «Мир», 2004.
2. Ролан Ж.-К., Селоши А., Селоши Д. Атлас по биологии клетки.
М., 2008.
3. Ченцов Ю.С. Введение в клеточную биологию. М., 2004.
4. Заварзин А.А., Харазова А.Д. Основы общей цитологии. – Л., 1982.
5. Ченцов Ю.С. Основы цитологии. – М., 1984.
ПЛАН ПРОВЕДЕНИЯ ЛАБОРАТОРНЫХ ЗАНЯТИЙ
Методические рекомендации к выполнению лабораторных занятий: прежде чем приступить к работе, нужно осмыслить значение и цель работы, внимательно прочитать и уяснить, что нужно делать, как ее оформить, затем – изучить теоретический материал по рекомендуемой литературе. Выполните задания, опишите ход эксперимента и сделайте соответствующие выводы. И в конце работы приводится ответы на контрольные вопросы.
ГОУ ВПО «ВЛАДИВОСТОКСКИЙ ГОСУДАРСТВЕННЫЙ
МЕДИЦИНСКИЙ УНИВЕРСИТЕТ РОСЗДРАВА»
ПАТОЛОГИЯ КЛЕТКИ
Учебное пособие
Владивосток – 2008
Утверждено РИСО ученого совета Владивостокского государственного медицинского университета
Рецензенты:
Д. б.н., профессор, заведующий кафедрой клеточной биологии ДВГУ
Д. м.н., профессор, заведующий кафедрой патофизиологии с курсом клинической патофизиологии ОГМА
Д. м.н., профессор, заведующий кафедрой патофизиологии ДВГМУ.
В пособии представлены современные сведения о причинах и механизмах развития структурно-функциональных нарушений клетки на молекулярном уровне. Рассмотрены вопросы о повреждении плазмолеммы клетки и ее внутренних структур: ядра, митохондрий, лизосом, эндоплазматического ретикулума и др., дана их клинико-патофизиологическая оценка. Показана роль изменения информационного обеспечения клетки в ее патологии. Отражено значение защитно-приспособительных механизмов. Отдельный раздел посвящен основам этиотропной и патогенетической профилактики и терапии патологии клетки.
Пособие рассчитано на студентов медицинских вузов, может быть использовано в процессе обучения интернов и ординаторов. Представляет интерес для аспирантов и врачей различных специальностей.
Введение | ||
Общая характеристика реакции клетки на повреждение | ||
Определение понятия «Патология клетки»: причины и виды повреждения | ||
Патология клеточной мембраны | ||
Патология клеточных органелл | ||
4.1. Патология ядра | ||
4.2. Патология митохондрий | ||
4.3. Патология лизосом | ||
4.4. Патология пероксисом | ||
4.5. Патология эндоплазматического ретикулума | ||
4.6. Патология комплекса аппарата Гольджи (пластинчатый комплекс) | ||
4.7. Патология цитоскелета | ||
Характерные изменения в цитозоле. | ||
Нарушение информационных процессов в клетке | ||
Механизмы адаптации клетки | ||
Некроз и апоптоз клетки | ||
Основы этиотропной и патогенетической профилактики и терапии патологии клетки | ||
Задание для самоконтроля | ||
Эталоны ответов | ||
Литература | ||
Введение
Человеческий организм состоит из 107 клонов клеток, включающих около 60 триллионов единиц. Выполнение работы того или иного органа в конечном счете определяется структурно-функциональным состоянием данных элементарных образований. Взаимодействие реагирующих систем организма с этиологическими факторами, особенно на фоне неблагоприятных условий, повреждают клетку: нарушается функция органов и систем – развивается болезнь.
В последнее время, благодаря стремительному развитию молекулярной биологии , цитологии , биофизики , биохимии и других смежных дисциплин значительно расширились наши представления о функции клетки и возможности ими управлять. Проект «Геном» и технология рекомбинантной ДНК углубили представления о генетических нарушениях. Появилась реальная возможность введения генов в клетку для исправления поврежденных наследственных механизмов. Прояснились многие вопросы о сложных сигнальных системах и взаимодействиях, регулирующих рост, развитие и программируемую смерть клетки (апоптоз).
Все эти достижения позволили переосмыслить возникновение, развитие и исходы многих заболеваний на качественно новом уровне – клеточно-молекулярном . Понимание механизмов развития патологических процессов и болезней на данном уровне, сделало реальным осуществлять молекулярную диагностику и прогнозирование течения для многих из них. Особенно это успешно применяются при таких заболеваниях как рак, диабет, болезни сердца, генетических дефектах и др. Назначение лекарственных препаратов с учетом путей их доставки в пораженную клетку способствует лечению больного на клеточно-молекулярном уровне, что значительно повышает процент их исцеления.
Несомненно, данный раздел медицины и впредь будет развиваться успешно (в первую очередь, учитывая быстрое прогрессирование молекулярной биологии). По утверждению многих исследователей «молекулярной медицине » принадлежит будущее. Поэтому изучение общих закономерностей возникновения, развития и исходов патологических процессов и заболеваний на этом уровне необходимы врачам всех специальностей.
1. ОБЩАЯ ХАРАКТЕРИСТИКА РЕАКЦИИ КЛЕТКИ НА ПОВРЕЖДЕНИЕ
Клетка является целостной биологической системой, имеющей все проявления жизни в сложном организме. Ее функционирование представляет собой реализацию определенных генетических программ, активность или пассивность последних определяется влиянием внешней среды, микроокружением (и внутренними потребностями клетки). В многоклеточном организме это контролируется различными местными и системными биорегуляторами.
Нормальная клетка, находящаяся в покое, поддерживает оптимальный метаболический уровень, соответствующий условиям (требованиям) среды. Данное стабильно-динамическое состояния получило название гомеостаза , гомеостазиса (от греч. homos – один и тот же, одинаковый + statis – стояние, неподвижность).
При взаимодействии клетки с различными физиологическими стимулами происходит переход на новый, чаще более активный уровень жизнедеятельности, без снижения ее жизнедеятельности. При этом, параметры гомеостаза не выходят за пределы максимально допустимых значений, а метаболическая активность и функциональные возможности клетки могут значительно повышаться. Такой ответ называется адаптацией клетки (от лат. adaptatio – приспособление). Возрастает, или наоборот, снижается активность действующих программ, включаются новые, раннее репрессированные, что приводит к увеличению синтеза ферментов, белков, вплоть до увеличения количества клеточных органоидов и их гипертрофии (увеличения массыферментов, белков, вплоть до увеличения количества клеточных органоидов и их гипертрофии ()ению синтезаность и функциональные в).
Таким образом, включая механизмы адаптации (а это не что иное, как комплекс защитно-приспособительных реакций) клетка может длительное время выполнять свою функцию в полном объеме, но в уже изменившихся условиях.
Взаимодействие клетки с патогенным агентом всегда сопровождается изменением ее стабильно-динамического состояния (гомеостаза), что очень часто проявляется различными структурными и функциональными нарушениями. Наличие последних, степень их выраженности, во многом определяется «мощностью» защитно-приспособительных механизмов, в задачу которых входят (как и при ответе на физиологический раздражитель) нормализация клеточного гомеостаза и обеспечение ее адекватного функционирования в изменившихся условиях. Принципиальных различий между механизмами адаптации на физиологический или патогенный раздражитель нет.
В связи с этим можно наблюдать следующие ситуации (рис. 1):
· активность (мощность) механизмов адаптации клетки такова, что они с успехом нивелируют негативные эффекты патогенного агента (достаточные резервы). Это обеспечивает работу клетки на новом, стабильно-динамическом уровне, при этом параметры гомеостаза не выходят за предельно допустимые значения, и повреждение клетки не происходит. После прекращения действия патогенного агента клетка может вернуться к исходному состоянию;
· адаптационные возможности клетки недостаточны (недостаточны резервы) для полного устранения действия патогенного агента. Параметры гомеостаза не могут сохраняться в пределах нормы, что и проявляется в виде различных видов патологии клетки. Процесс, предшествующий гибели клетки, и представляющий собой начальные, обратимые стадии ее повреждения, получил название паракнекроз (от грeч. para – пара, при + nekros – мертвый);
· по мере истощения защитно-приспособительных механизмов или в силу их несовершенства (они сами могут порождать вторичные нарушения «эндогенезация патологического процесса», 2004), клетка подходит к следующему этапу ответа на патогенный агент – некробиоз (от греч. nekros – мертвый, bios – жизнь). Это глубокая, частично не обратимая стадия повреждения клетки, непосредственно предшествующая моменту ее смерти.
 повреждающий
достаточные
повреждающий
достаточные
https://pandia.ru/text/78/044/images/image015_18.gif" width="87"> агент резервы
 |  |
недостаточные - гипоксический
резервы - свободнорадикальный
https://pandia.ru/text/78/044/images/image020_17.gif" width="30" height="74">.gif" height="50 src="> точка
необратимости
https://pandia.ru/text/78/044/images/image031_11.gif" width="14" height="14"> паранекроз некробиоз
https://pandia.ru/text/78/044/images/image034_8.gif" width="18" height="144">
https://pandia.ru/text/78/044/images/image037_8.gif" width="143" height="12">![]()
![]()
https://pandia.ru/text/78/044/images/image043_8.gif" width="386"> - неспособность к самостоятельной энергопродукции
https://pandia.ru/text/78/044/images/image044_8.gif" width="386"> - эндогенный детергентный эффект
https://pandia.ru/text/78/044/images/image047_5.gif" width="115"> коагуляционный, колликвационный , казеозный некроз
(цепь посмертных
апоптоз изменений в клетке)
Рис. 1. Различные варианты ответа клетки на повреждение.
К – клетка. В овале – максимальные границы нормы показателей гемостаза клетки, пунктирная - возвращение к исходному состоянию(обратимость ответа)
Значительно реже, вместо слова некробиоз, используют термин «преднекроз ». Самый негативный исход ответа клетки на патогенный агент является некроз – посмертные изменения необратимого характера, характеризующиеся ферментативным разрушением клеточных структур и денатурацией ее белков. Как правило, некроз наступает вслед за некробиозом. Между двумя этими вида ответа существует точка необратимости , пройдя которую, клетка утрачивает всякую возможность остаться живой. Цитологическими критериями необратимости считаются изменения ядра – его конденсация (кариопикноз ), с последующим распадом на конденсированные глыбки (кариорексис ) или растворением (кариолизис ). По биохимическим критериям клетка считается погибшей с момента полного прекращения синтеза АТФ .
При взаимодействии клетки с этиологическим фактором необычным по своей природе, или чрезвычайного, экстремального характера, этапности ответа на них может не наблюдаеться, в ней практичности сразу развивается цепь посмертных изменений, т. е. некроз. Объясняется это следующим: на необычные по своей природе патогенные агенты, в процессе эволюции в организме не сформировались механизмы защиты от него (они ранее не встречалисьэволюции в организме не сформировались механизмы защиты ()нты, в процессектера ()нтеза АТФ. атологии клетки. за преде) или мощность экстремальных факторов значительно превышает возможности адаптации клетки.
Существует и другой вариант гибели клетки – апоптоз (от греч. apo – отделение, удаление + ptosis – падение). Поэтический перевод – «опадание листьев ». Апоптоз определяют как программированную клеточную смерть, в его развитии активную роль играют специализированные и генетически запрограммированные внутриклеточные механизмы.
Рассмотренные варианты ответа клетки на патогенный агент мы используем в качестве своеобразного плана изложения учебного материала. По нашему мнению, это не только улучшит восприятие различных звеньев патогенеза, но и будет способствовать более глубокому пониманию динамической взаимосвязи между ними.
Начнем с определения понятия «патология клетки» и характеристики этиологических факторов вызывающих ее повреждение.
2. ОПРЕДЕЛЕНИЕ ПОНЯТИЯ «ПАТОЛОГИЯ КЛЕТКИ»: ПРИЧИНЫ И ВИДЫ ПОВРЕЖДЕНИЯ.
Патология клетки – типовой патологический процесс, характеризующийся нарушением внутриклеточного гомеостаза, что ограничивает функциональные возможности клетки и может приводить ее к гибели или снижению продолжительности жизни.
Гомеостаз клетки – способность клетки существовать при изменении условий обитания с сохранением устойчивого динамического равновесия со средой.
Понятие «гомеостаз клетки » включает в себя ряд показателей (констант): внутриклеточное постоянство ионов водорода , электронов, кислорода, субстратов для энергетического и пластического обеспечения жизнедеятельности клетки, ферментов, нуклеотидов и еще ряд веществ.
Константы (лат. constantus – постоянная величина) гомеостаза клетки зависят от:
· структурно-функционального состояния ее различных мембран (плазмолемы, митохондрий, лизосом и др.) и органелл, интенсивности течения внутриклеточных биохимических процессов. Это своеобразная «метаболическая составляющая гомеостаза » и определяется работой исполнительного аппарата клетки;
· информационных процессов . Нормальная жизнедеятельность клетки невозможна без информации, поступающей к ней из внешней среды. Очень часто она изменяет параметры внутриклеточного постоянства, что является следствием включения приспособительных (адаптивных) программ, позволяющих клетке оптимально приспосабливаться к конкретной ситуации согласно поступившей информации. «Правильность » изменения констант внутриклеточного гомеостаза и их поддержание в границах нормы в данном случае определяется в первую очередь количеством и качеством информационного обеспечения клетки (наличием сигнальных молекул, рецепторов, пострецепторных связей и др.). Исполнительный аппарат клетки выполняет лишь «полученные указание ».
Следовательно, патология клетки может возникнуть и без первичного «полома » ее исполнительного аппарата, а из-за нарушений в механизмах сигнализации, в так называемой «информационной составляющей » внутриклеточного гомеостаза.
В зависимости от природы этиологического фактора, нарушающего гомеостаз (метаболическое и/или информационное его составляющее) клетки, различают физические, химические и биологические повреждающие агенты.
Физические этиологические факторы – это механические и температурные воздействия (гипо - и гипертермия), энергия электрического тока, ионизирующей радиации и электромагнитных волн, влияние факторов космического полета (ускорение, гипокенезия) и др.
Химические этиологические факторы – воздействие многочисленных неорганических и органических веществ (кислоты, щелочи, соли тяжелых металлов, этиловый и метиловый спирт). Патология может быть обусловлена дефицитом или избытком белков, жиров, углеводов, витаминов , микроэлементов и др. веществ. Немаловажное значение в этой группе факторов имеют и лекарственные препараты.
Все, выше названные патогенные факторы, вызывают различные повреждения клеток.
Тип (вид) повреждения клетки зависит от :
· скорости развития основных проявлений нарушений функции клеток . Выделяют острое и хроническое повреждение клетки. Острое повреждение развивается быстро, и как правило, в результате однократного, но интенсивного повреждающего воздействия. Хроническое повреждение протекает медленно и является следствием многократного влияния, но менее интенсивного по силе повреждения агента;
· жизненного цикла клетки, на период которого приходится воздействие повреждающего фактора. Различают митотические и интерфазные повреждения;
· от степени (глубины) нарушения клеточного гомеостаза – обратимые и необратимые повреждения;
· от характера взаимодействия повреждающего фактора с клеткой . Если патогенный агент действует непосредственно на клетку, то говорят о прямом (первичном) ее повреждении. В условиях целостного организма влияние причины может осуществляться и через формирование цепи вторичных реакций. Например, при механической травме непосредственно в месте воздействия этого агента образуются биологически активные вещества (БАВ) – это продукты распада погибших клеток, гистамин, оксидазы, простогландины и др. соединения, синтезируемые поврежденными клетками. БАВ, в свою очередь, вызывают нарушения функции клеток, ранее не попавших под влияние данного фактора. Такое повреждение получило название опосредованное или вторичное . Воздействие этиологического фактора может проявляться опосредованно и через изменения нервных и эндокринных регуляций (шок, стресс), при отклонениях физико-химического состояния организма (ацидоз, алколоз), при нарушениях системного кровообращения (сердечная недостаточность), гипоксии, гипо - и гипертермия, гипо - и гипегликемия и др.
· от характера повреждений вызванных определенным патогенным фактором . Рассматривают специфические и неспецифические повреждения.
(2002) выделяет и специфические повреждения определенных клеток, возникающее при взаимодействии с самыми различными патогенными факторами. В качестве примера приводит развитие контрактур мышечных клеток при влиянии на них физических, химических и биологических факторов, или, возникновение гемолиза эритроцитов при аналогичных воздействиях.
Неспецифические повреждения – это стандартные, стереотипные изменения в клетках возникающие при их взаимодействии с широким спектром этиологических факторов. В качестве примера можно привести следующие нарушения:
· повышение проницаемости мембран клеток;
· внутриклеточный ацидоз;
· денатурация молекул белков;
· дисбаланс ионов и воды;
· изменение интенсивности окислительного фосфорилирования.
Взаимосвязи между специфическими и неспецифическими повреждениями клеток разнообразны. Они могут возникать одновременно, либо одно из них предшествует другому. Выяснение конкретных видов нарушений, времени их возникновения и соотношении между собой, дает врачу необходимую информацию о характере и интенсивности действия причинного фактора, глубине и распространенности патологического процесса. Это в свою очередь обеспечивает проведение более этиотропной и патогенетической профилактики и терапии. Например, если при гепатитах различного происхождения регистрируется только увеличение в плазме крови концентрации ионов К и аланинаминотрансферазы (АЛТ) то это свидетельствуют о легком течении или начале заболевания. Калий и АЛТ находятся в цитоплазме, возрастание их содержание за пределами клеточной мембраны характерны при нарушении ее проницаемости (неспецифическое повреждение). Появление же в крови довольно специфического для печени фермента – сорбитдегидрогеназы и органеллоспецифичных – глютаматдегидрогеназы (локализация - митохондрии), кислой фосфотазы (локализация лизосомы) говорит об усугублении патологического процесса . Он уже не ограничивается только мембраной клетки, а затрагивает и внутриклеточные структуры.
Как было отмечено выше, патология клетки возникает вследствие нарушения ее гемостаза. Он может изменяться не только при непосредственном воздействии патогенного агента на клетку (тем самым, нарушая в основном работу ее исполнительного аппарата), но и при недостаточности информационных механизмов, инициирующих включение тех или иных адаптогенных программ. В связи с этим, природу заболеваний человека можно рассматривать с двояких позиций – материально-энергетических и информационных (, 1999). Болезнь развивается, и при повреждении исполнительного аппарата клетки (материально-энергетическая позиция ), и при нарушении ее информационных механизмов (информационная позиция ). Основываясь на последнем положеним, существует даже специальная терминология – «болезни регуляции », «дизрегуляционная патология ».
Данные позиции легче выявляются на начальных этапах патологии клетки. По мере ее развития различия между ними более затруднительны, и тем не менее, этиотропная и патогенетическая терапия будет более адекватной и успешной при установлении истинного механизма (причины) развития того или иного проявления патологии.
Сейчас мы приступаем непосредственно к рассмотрению ответа клетки на патогенный агент. Согласно нашего плана (рис. 1), сюда входят вопросы адаптации и паранекроза. Они между собой тесно связаны, так как любой патологический процесс (болезнь) состоит из двух компонентов: повреждения (альтерации ) и защитно-приспособительных (адаптивных ) механизмов. Альтерация моментально вызывает активацию адаптивных механизмов, направленных на поддержание жизнедеятельности клетки в изменившихся условиях. Параллельное изучение вопросов альтерации и защитно-приспособи-тельных механизмов создает определенные трудности в усвоении учебного материала. Поэтому мы первоначально разберем механизмы повреждения гомеостаза клетки, а затем защитно-приспособительных реакций. При этом будем помнить, что начальный этап альтерации клетки – паранекроз – это не только повреждение, но и наличие защитно-приспособительных механизмов, пусть и не в полной мере выполняющих свое назначение.
Рассмотрение патологии клетки начинаем с нарушений, возникающих при непосредственном воздействии на нее патогенного агента. Взаимодействие этиологического фактора с различными структурными образованиями клетки, ведет к нарушению ее гомеостаза (его метаболической составляющей ), и, следовательно, развитию болезни. Патология может возникнуть при повреждении различных биомембран клетки (особенно часто повреждается плазмолемма) и внутриклеточных образований: ядра, митохондрий, лизосом и др. (рис. 2).
3. ПАТОЛОГИЯ КЛЕТОЧНОЙ МЕМБРАНЫ.
Плазматической мембраной называется барьер, который окружает цитоплазму, определяя границы клетки. Ей присущи следующие функции:
· контроль состава внутриклеточной среды;
· обеспечение и облегчение межклеточной и внутриклеточной информации;
· обеспечение образования тканей с помощью межклеточных контактов.
Современная жидкостно-мозаичная модель мембраны представляет плазмолемму как бы «море» липидов с белковыми «айсбергами » и «островами» (рис. 3). Основу мембраны составляет двойной слой амфифильных липидов, причем, каждая молекула мембранного липида имеет гидрофильную головку – 1, рис. 3 (растворяется в воде) и два гидрофобных хвоста – 2, рис. 3 (в воде не растворяются). Отсюда и термин – «амфифильность» – он обозначает молекулы, имеющие в своем составе как водо - так и водонерастворимые группы.
В водной среде такие соединения самопроизвольно образуют бислой, в котором гидрофобные части молекул ориентированы друг к другу, а гидрофильные – к воде. К основным мембранным липидам относят: фосфолипиды, сфинголипиды, гликолипиды и холестерин – 7, рис. 3.


shortcodes">
1
5.
В медико-генетическую консультацию обратилась женщина 20 лет. Ее родная сестра больна тяжелой формой серповидно-клеточной анемии, у пациентки никаких заболеваний крови не было, супруг здоров. Женщину интересует, каков риск развития этой болезни у планируемого ребенка. При обследовании крови супругов на типы гемоглобина получены результаты: у мужчины HbA 98 %, HbS 1 %; у женщины HbA 70 %, HbS 29 %.
Каков ответ на вопрос женщины? Были ли основания для беспокойства? Возможна ли профилактика при планировании конкретного ребенка? Связано ли заболевание с полом ребенка?
6.
Какие группы крови невозможны у детей от родителей со следующими группами крови по системе АВ0: I(0) и III(В)? III(В) и IV(АВ)? IV(АВ) и IV(АВ)? II(А) и III(В)? Какое значение при рождении второго ребенка имеет установленная группа крови первого?
7.
В медико-генетическую консультацию обратилась беременная, которая сообщила, что ее сестра больна фенилкетонурией, сама наследственные заболевания отрицает. Супруг здоров. В его роду были браки между близкими родственниками, но случаев фенилкетонурии не отмечалось.
Какова вероятность появления фенилкетонурии у ребенка? Имеет ли значение вероятный пол ребенка? Можно ли лечить эту болезнь после ее появления?
Глава 4
ПАТОЛОГИЯ КЛЕТКИ
Клетка – структурная и функциональная единица всех живых организмов.
В клетке сосредоточено уникальное свойство живого – способность размножаться, видоизменяться и реагировать на изменения окружающей среды. Эукариотическая клетка состоит из трех основных компонентов: плазматической мембраны, ядра, цитоплазмы. Главной функцией клетки является осуществление обмена со средой веществом, энергией и информацией, что подчинено в конечном счете задаче сохранения клетки как целого при изменении условий существования (рис. 4.1 на с. 52).
Органоиды клетки, обладая определенными морфологическими особенностями, обеспечивают основные проявления жизнедеятельности клетки. С ними связаны дыхание и энергетические запасы (митохондрии), синтез белков (рибосомы, шероховатая эндоплазматическая сеть), накопление и транспорт липидов и гликогена, обезвреживание токсинов (гладкая эндоплазматическая сеть), синтез продуктов и их выделение из клетки (комплекс Гольджи), внутриклеточное пищеварение и защитная функция (лизосомы). Важно подчеркнуть, что функции субклеточных органелл не строго разграничены, поэтому они могут участвовать в разных внутриклеточных процессах.
Все перечисленное делает познание основ патологии клетки абсолютно необходимым для понимания закономерностей развития патологии на уровне тканей, органов и систем, болезни в целом – на уровне организма человека.
Рис. 4.1.
Общее строение эукариотической клетки и ее основных органелл :
1
– секреторные гранулы (накопление продуктов секреции); 2
– центриоли (центр полимеризации микротрубочек); 3
– гладкая эндоплазматическая сеть (детоксикация и синтез стероидов); 4
– лизосомы (внутриклеточное переваривание); 5
– митохондрия (синтез АТФ и стероидов); 6
– сферические единицы (превращение энергии); 7
– липидные капельки (накопление); 8
– ядрышко (синтез рРНК); 9
– ядерная оболочка (разделение хроматина и цитоплазмы); 10
– шероховатая эндоплазматическая сеть (синтез и сегрегация белков, посттрансляционные изменения); 11
– комплекс Гольджи (конечные посттрансляционные изменения, упаковка и транспорт)
4.1. ПОВРЕЖДЕНИЕ КЛЕТКИ: ПРИЧИНЫ И ОБЩИЕ МЕХАНИЗМЫ
Повреждение – процесс, проявляющийся нарушением структурной и функциональной организации живой системы, вызванный различными причинами.
В наиболее общем смысле повреждение на любом уровне представляет собой такое изменение структуры и функции, которое не способствует, а мешает жизни и существованию организма в окружающей среде. Повреждение является начальным моментом в развитии патологии, внутренней стороной взаимодействия причинного фактора с организмом. В этом смысле термины «этиологический фактор», «болезнетворный фактор» и «повреждающий фактор» являются синонимами.
Любое повреждение проявляется на различных уровнях:
молекулярном (повреждение клеточных рецепторов, молекул ферментов, нуклеиновых кислот вплоть до их дезинтеграции);
субклеточном – ультраструктурном (повреждение митохондрий, эндоплазматической сети, мембран и других ультраструктур вплоть до их деструкции);
клеточном (различные дистрофии из-за нарушения разных видов обмена с возможным развитием некроза по типу рексиса или лизиса клетки);
тканевом и органном (дистрофические изменения в большинстве клеток и строме с возможным развитием некроза (по типу инфаркта, секвестра и др.);
организменном (болезнь с возможным смертельным исходом).
Иногда дополнительно выделяют уровень тканевых комплексов, или гистионов, включающих в свой состав сосуды микроциркуляторного русла (артериола, капилляры, венула) и питаемые ими клетки паренхимы, соединительную ткань и терминальные нервные окончания. Морфологически повреждение может быть представлено двумя патологическими процессами: дистрофией и некрозом, которые нередко являются последовательными стадиями (рис. 4.2).
Причины повреждения клетки. Вовлечение клеток во все патологические процессы, происходящие в организме, объясняет и универсальность причин, вызывающих повреждение клеток, которые соответствуют по структуре классификации этиологических факторов болезни вообще (табл. 4.1).
А – нормальная клетка: 1 – ядро; 2 – лизосома; 3 – эндоплазмолитическая сеть; 4 – митохондрии.
Б – обратимое повреждение: 1 – объединение внутримембранных частиц;
2 – разбухание эдоплазматической сети;
3 – дисперсия рибосом; 4 – разбухание митохондрий; 5 – уменьшение плотности митохондрий; 6 – самопереваривание лизосом; 7 – агрегация ядерного хроматина; 8 – выпячивание.
В – необратимые повреждения: 1 – миелиновые тельца; 2 – лизис эндоплазматической сети; 3 – дефект клеточной мембраны; 4 – большая разряженность митохондрий; 5 – пикноз ядра; 6 – разрыв лизосом и аутолиз
Причиной повреждения клетки может стать фактор как экзогенной, так и эндогенной природы. Применительно к клетке наиболее важные механические и физические агенты (механическая травма, колебания температуры окружающей среды и атмосферного давления, радиация, электрический ток, электромагнитные волны); химические агенты (изменение pH, снижение содержания кислорода, соли тяжелых металлов, органические растворители и др.); всевозможные инфекционные агенты; иммунные реакции, генетические нарушения, дисбаланс питания.
Таблица 4.1
Этиологические факторы повреждения клетки

Психогенные факторы повреждения для организма на уровне клеток воспринимаются через вторичные воздействия, которые являются физическими или химическими по своей природе. Например, при эмоциональном стрессе повреждение миокарда объясняется воздействием адреналина и изменением электрической активности симпатических волокон автономной нервной системы.
Общий патогенез клеточного повреждения.
С точки зрения развития процессов в самой общей форме повреждения клетки могут проявляться нарушениями клеточного обмена веществ, развитием дистрофии, парабиоза и, наконец, некроза, когда клетка погибает.
Повреждения клетки могут быть обратимыми
и необратимыми
. Например, обратимым является повреждение лизосом в клетках эпителия кишечника под влиянием эндотоксина микроорганизмов кишечной группы. После прекращения интоксикации лизосомы в поврежденной клетке восстанавливаются. В случае повреждения клеток энтеровирусом повреждение выражается дегрануляцией лизосом, которую может вызвать, например, любая вирусная инфекция.
По своему течению повреждения могут быть острыми
и хроническими
. Функциональные проявления острого повреждения клетки делятся на преддепрессионную гиперактивность, парциальный некроз и тотальное повреждение (клеточный некроз).
Первое и наиболее общее неспецифическое выражение повреждения клетки при действии любого агента – это нарушение состояния неустойчивого равновесия клетки и среды, являющегося общей характеристикой всего живого, независимо от уровня его организации.
Преддепрессионная гиперактивность (по Ф. З. Меерсону) возникает вследствие обратимого повреждения клетки умеренными воздействиями патогенных факторов. В результате в мембране клетки происходят неспецифическое возбуждение и усиление деятельности органелл, в первую очередь митохондрий. Это приводит к усилению окисления субстратов и синтеза АТФ, сопровождается повышением резистентности клетки к патологическому фактору. Если воздействие этого фактора ограничено, может произойти ликвидация повреждения с последующим восстановлением первоначальной структуры и функции. Считают, что после такого воздействия в генетическом аппарате клетки сохраняется информация о происшедшем воздействии, так что в дальнейшем при повторном действии этого же фактора приспособление клетки значительно облегчается.
В случае парциального некроза поврежденная часть клетки отделяется от функционирующей части вновь образующейся мембраной и уничтожается фагоцитами. После этого структура и функция клетки восстанавливаются за счет гиперплазии субклеточных единиц.
Если повреждающий фактор имеет выраженную интенсивность и время действия, то происходит тотальное повреждение клетки, что приводит к прекращению функции митохондрий, нарушению клеточного транспорта и всех энергозависимых процессов. В дальнейшем происходит массивное разрушение лизосом, выход гидролитических ферментов в цитоплазму и расплавление остальных органелл, ядра и мембран. Фаза острого повреждения клетки, когда еще сохраняется небольшой градиент концентрации ионов между цитоплазмой и внеклеточной средой, называется агонией клетки. Она необратима и завершается некрозом клетки, при этом резкое увеличение проницаемости и частичное разрушение клеточных мембран способствуют доступу в клетку из окружающей среды ферментов, которые продолжают разрушение всех ее структурных элементов.
Специфическое и неспецифическое в повреждении клетки.
Специфические повреждения можно усмотреть при анализе любого его вида. Например, при механической травме – это нарушение целостности структуры ткани, при аутоиммунной гемолитической анемии – изменение свойств мембраны эритроцитов под влиянием гемолизина и комплемента, при радиационном повреждении – образование свободных радикалов с последующим нарушением окислительных процессов.
Неспецифическими повреждениями
клетки, т. е. мало зависящими от вида повреждающего фактора, являются следующие:
нарушение неравновесного состояния клетки и внешней среды;
нарушение структуры и функции мембран: проницаемости и мембранного транспорта, мембранного электрического потенциала, рецепторного аппарата, формы клеток;
нарушение обмена и электролитного состава клетки и ее отдельных частей;
нарушение активности ферментных систем клетки (вплоть до ферментативного разрушения клетки);
уменьшение объема и интенсивности биологического окисления;
нарушение хранения и передачи генетической информации;
снижение специфической функции (для специализированных клеток).
Повреждение специфических функций, нужных для организма в целом, прямо не отражается на судьбе клеток, но определяет суть изменений в органах и системах, поэтому рассматривается в курсе частной патологии.
Большинство повреждений на субклеточном уровне имеет неспецифический характер и не зависит от вида повреждающих факторов. Так, например, в миокарде при острой ишемии, воздействии адреналина, отравлении морфином, разлитом гнойном перитоните, облучении наблюдаются аналогичные изменения поврежденных клеток в виде набухания митохондрий и разрушения их мембран, вакуолизации эндоплазматической сети, очаговой деструкции миофибрилл и появления избыточного количества липидных включений. Такие идентичные изменения структур под влиянием различных факторов называются стереотипными.
При одинаковом воздействии на весь орган какого-либо повреждающего фактора обычно проявляется весь спектр возможных состояний клетки от практически нормального и даже усиленно функционирующего до гибели (некроза). Это явление называется мозаичностью
. Например, при действии вируса ветряной оспы на клетки кожи некрозы развиваются в виде мелких очагов, образуя характерную сыпь в виде пузырьков (везикул).
Повреждения на клеточном уровне иногда могут иметь специфический характер. Специфические изменения обусловлены внутриклеточной репликацией вируса (с появлением в ядре или цитоплазме включений, представляющих собой или скопления вирусных частиц, или реактивные изменения клеточного вещества в ответ на их репликацию), опухолевым метаморфозом и врожденными или приобретенными ферментопатиями, приводящими к накоплению в клетке нормальных метаболитов в избыточном количестве или аномальных в виде включений.
4.2. ПАТОЛОГИЯ КЛЕТОЧНЫХ МЕМБРАН
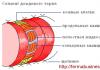
Основной структурной частью мембраны является липидный бислой, состоящий из фосфолипидов и холестерина с включенными в него молекулами разных белков. Снаружи клеточная мембрана покрыта слоем гликопротеидов. К функциям мембраны клетки относятся избирательная проницаемость, реакции межклеточных взаимодействий, поглощение и выделение специфических веществ (рецепция и секреция). Плазматическая мембрана – место приложения физических, химических, механических раздражителей внешней среды и сигналов информационного характера из внутренней среды организма. Информационная функция обеспечивается рецепторами мембраны, защитная – самой мембраной, контактная – клеточными стыками (рис. 4.3).
Способность формировать мембраны является решающей в образовании клетки и ее субклеточных органелл. Любое нарушение сопровождается изменением проницаемости клеточных мембран и состояния цитоплазмы поврежденной клетки. Повреждение клеточных мембран может быть обусловлено деструкцией их липидных или белковых (ферментных и рецепторных) компонентов.
К патологии клетки могут вести нарушения следующих функций мембран: мембранного транспорта, проницаемости мембран, коммуникации клеток и их «узнавания», подвижности мембран и формы клеток, синтеза и обмена мембран (схема 4.1).

1 –двойной слой фосфолипидов; 2 – мембраные белки; 3 – полисахаридные цепи
Схема 4.1. Общие механизмы повреждения мембран клеток [Литвицкий П. Ф. , 1995]

Повреждение липидных компонентов клеточных и субклеточных мембран возникает несколькими путями. Важнейшими из них являются перекисное окисление липидов, активация мембранных фосфолипаз, осмотическое растяжение белковой основы мембран, повреждающее воздействие иммунных комплексов.
Мембранный транспорт предполагает перенос ионов и других субстратов против избытка (градиента) их концентрации. При этом нарушаются функция клеточных насосов и процессы регуляции обмена веществ между клеткой и окружающей ее средой.
Энергетической основой работы клеточных насосов являются процессы, зависящие от энергии АТФ. Эти ферменты «вмонтированы» в белковую часть клеточных мембран. В зависимости от вида проходящих по каналу ионов различают Na – K-АТФазу, Ca – Mg-АТФазу, Н – АТФазу и др. Особое значение имеет работа первого насоса, результатом которой является превышение концентрации К + внутри клетки приблизительно в 20–30 раз по сравнению с внеклеточной. Соответственно этому концентрация Na + внутри клетки приблизительно в 10 раз меньше, чем снаружи.
Повреждение Na – K-насоса вызывает освобождение К + из клетки и накопление в ней Na + , что характерно для гипоксии, инфекционных поражений, аллергии, снижения температуры тела и многих других патологических состояний. С транспортом Na + и К + тесно связан транспорт Ca 2+ . Интегральное выражение этих нарушений хорошо иллюстрируется на примере гипоксии миокарда, которая проявляется прежде всего патологией митохондрий.
Известно участие Са 2+ в освобождении медиаторов аллергии из лабиринтов (тучных клеток). По современным данным, их аллергическая травма сопровождается разжижением мембраны, разрыхлением и увеличением проводимости кальциевых каналов. Ионы кальция, проникая в большом количестве внутрь клетки, способствуют освобождению гистамина и других медиаторов из гранул.
Морфологически нарушение проницаемости плазматической мембраны проявляется усиленным образованием ультрамикроскопических пузырьков, что приводит к дефициту поверхности или, напротив, увеличению поверхности за счет мембран микропиноцитозных пузырьков. В отдельных случаях выявляются утолщение и извитость участков мембраны, отделение части цитоплазмы, окруженной мембраной, от клетки. Это свидетельствует об активизации цитоплазматической мембраны. Другим наблюдаемым при электронной микроскопии признаком повреждения мембраны является образование крупных микропор – «брешей», что ведет к набуханию клетки, перерастяжению и разрыву клеточных мембран.
С формой и подвижностью мембраны непосредственно связаны изменения формы и подвижности клетки в целом, хотя при патологии обычно происходит упрощение формы клеточной поверхности (например, потеря микроворсинок энтероцитами).
Отдельного внимания заслуживает патология, развивающаяся при повреждении межклеточных взаимодействий. Поверхность мембраны клетки содержит множество рецепторов, воспринимающих различные раздражители. Рецепторы представлены сложными белками (гликопротеидами), способными свободно перемещаться как по поверхности клеточной мембраны, так и внутри ее. Механизм рецепции является энергозависимым, поскольку для передачи сигнала с поверхности внутрь клетки требуется АТФ. Особый интерес представляют рецепторы, одновременно являющиеся поверхностными антигенами-маркерами определенных типов клеток.
При разных патологических процессах (воспаление, регенерация, опухолевый рост) могут изменяться поверхностные антигены, причем различия могут касаться как типа антигена, так и его доступности со стороны внеклеточного пространства. Например, повреждения гликолипидов мембраны делают ее более доступной воздействию антител.
Патология клеточной рецепции ведет к нарушению восприятия информации. Например, наследственное отсутствие апо-Е– и апо-В-рецепторов у клеток печени и жировой клетчатки ведет к развитию семейных типов ожирения и гиперлипопротеинемии. Аналогичные дефекты выявлены при некоторых формах сахарного диабета.
Межклеточное взаимодействие и кооперация клеток определяются состоянием клеточных стыков, которые могут повреждаться при различных патологических состояниях и болезнях. Клеточные стыки выполняют три главные функции: межклеточную адгезию, «тесное общение» клеток и герметизацию слоя эпителиальных клеток. Межклеточная адгезия ослабевает при опухолевом росте уже на ранних этапах онкогенеза и является одним из критериев роста опухоли. «Тесное общение» заключается в прямом обмене клеток через щелевидные стыки информационными молекулами. Дефекты «тесного общения» играют значительную роль в поведении и возникновении злокачественных опухолей. Нарушения межмембранных связей клеток тканевых барьеров (кровь – мозг, кровь – легкие, кровь – желчь, кровь – почки) ведут к увеличению проницаемости плотных стыков клеток и повышенной проницаемости барьеров.
4.3. ПАТОЛОГИЯ КЛЕТОЧНОГО ЯДРА
Ядро обеспечивает координацию работы клетки в интерфазу, хранение генетической информации, передачу генетического материала при клеточном делении. В ядре происходят репликация ДНК и транскрипция РНК. При повреждении могут наблюдаться отек ядра, его сморщивание (пикноз), разрыв и разрушение (кариорексис и кариолизис). Ультрамикроскопическое исследование позволяет различить несколько типовых нарушений ядра и генетического аппарата клетки.
1. Изменение структуры и размеров ядра
зависит от содержания в нем ДНК. В нормальном интерфазном ядре содержится диплоидный (2n) набор хромосом. Если после окончания синтеза ДНК не происходит митоза, появляется полиплоидия – кратное увеличение набора ДНК. Полиплоидия может встречаться в нормально функционирующих клетках печени, почек, в миокарде; она особенно ярко выражена в тканях при регенерации и опухолевом росте, причем чем более злокачественна опухоль, тем более выражена гетероплоидия. Анеуплоидия – изменение в виде неполного набора хромосом – связана с хромосомными мутациями. Ее проявления встречаются в большом количестве в злокачественных опухолях.
Вещество ДНК в ядре распределено неравномерно. В наружных отделах ядер находят конденсированный хроматин (гетерохроматин), который считается неактивным, а в остальных отделах – неконденсированный (эухроматин), активный. Конденсация хроматина в ядре рассматривается как признак метаболической депрессии и предвестник гибели клетки. К патологическим изменениям ядра относят также его токсическое набухание. Уменьшение размеров ядра характерно для снижении обмена веществ в клетке и сопутствует ее атрофии.
2. Изменение формы ядра
может вызываться цитоплазматическими включениями (перстневидные клетки при слизьобразующем раке, ожиревшие гепатоциты), образованием множественных выпячиваний ядра в цитоплазму вследствие повышения синтетической активности ядра (полиморфизм ядер при воспалении, опухолевом росте). Как крайний вариант в ядре могут встречаться включения (цитоплазматические или вирусные).
3. Изменение количества ядер
проявляется многоядерностью в гигантских клетках при воспалении (клетки Пирогова–Лангханса при туберкулезе), опухолях (клетки Штернберга–Березовского при лимфогранулематозе). Безъядерность может наблюдаться в нормальных клетках (эритроциты, тромбоциты), в жизнеспособных фрагментах опухолевых клеток и как свидетельство гибели клеток (кариолизис).
4. Изменение структуры и размеров ядрышек
заключается в их увеличении и повышении плотности (соответствует повышению функциональной активности) или дезорганизации (встречается при энергодефиците в клетке и сопровождается патологией митозов).
5. Изменение ядерной оболочки (двойной мембраны)
состоит в нарушениях связи ее с эндоплазматической сетью, выпячивании и искривлении обеих мембран, изменении количества и размеров пор, появлению включений в межмембранном пространстве. Данные изменения свидетельствуют о вовлеченности ядра в повреждение клетки и характерны для интоксикации, вирусных инфекций, радиационных повреждений, опухолевого перерождения клетки.
6. Процессы клеточного деления (митоза)
могут нарушаться при различных воздействиях, при этом может страдать любое из его звеньев. Наибольшую известность получила классификация патологии митозов, предложенная И.А. Аловым (1972):
I тип
– повреждение хромосом (задержка деления в профазе);
II тип
– повреждение митотического аппарата (задержка в метафазе);
III тип
– нарушение цитотомии (задержка в телофазе).
Можно считать установленным, что задержка вступления клеток в митоз возникает в основном в связи с нарушением их метаболизма, в частности синтеза нуклеиновых кислот и белков, а нарушение хромосом при репродукции клетки, обнаруживаемое в условиях патологии, – вследствие разрыва цепей ДНК и расстройства репродукции ДНК хромосом.
Особенности реакции клетки на повреждающий фактор зависят как от его характеристики, так и от типа клетки по ее способности к делению, обеспечивающей возможность рекомпенсации. Считают, что в организме имеется три категории специализированных клеток по их способности к делению.
Клетки I категории
с самого рождения организма достигают высокоспециализированного состояния структур за счет минимизации функций. В организме отсутствует источник возобновления этих клеток в случае их дисфункции. К таким клеткам относятся нейроны. Клетки I категории способны к внутриклеточной регенерации, в результате которой восстанавливаются утраченные части клеток, если сохранены ядерный аппарат и трофическое обеспечение.
Клетки II категории
– это высокоспециализированные клетки, выполняющие какие-либо определенные функции и затем либо «изнашивающиеся», либо слущивающиеся с различных поверхностей, причем иногда очень быстро. Подобно клеткам I категории, они не способны размножаться, однако в организме имеется механизм для их непрерывного воспроизводства. Такие клеточные популяции называют обновляющимися, а состояние, в котором они находятся, – стационарным. К таким клеткам относятся, например, клетки, выстилающие большую часть кишечника.