Строение и функции ядра растительной клетки.
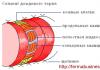
Ядро – обязательная часть эукариотической клетки. Это место хранения и воспроизведения наследственной информации. Ядро также служит центром управления обменом веществ и почти всех процессов, происходящих в клетке. Чаще всего в клетках имеется лишь одно ядро, редко - два или несколько. Форма его чаще всего шаровидная или эллипсоидальная. В молодых, особенно меристематических, клетках оно занимает центральное положение, но позднее обычно смещается к оболочке, оттесняемое растущей вакуолью. Снаружи ядро покрыто двойной мембраной – ядерной оболочкой, пронизанной порами (поры ядра - динамичные образования, они могут открываться и закрываться; таким путем может осуществляться регуляция обмена между ядром и цитоплазмой) на краях которых наружная мембрана переходит во внутреннюю. Наружная ядерная мембрана соединяется с мембранными каналами ЭПС. На ней располагаются рибосомы. Внутренняя мембрана может давать впячивания.
Внутреннее содержимое ядра – кариоплазма с погруженными в нее хроматином и ядрышками, и рибосомами. Кариоплазма (нуклеоплазма) представляет собой желеобразный раствор, который заполняет пространство между структурами ядра (хроматином и ядрышками). Она содержит ионы, нуклеотиды, ферменты.
Хроматин – это деспирализованная форма существования хромосом. В деспирализованном состоянии хроматин находится в ядре неделящейся клетке. Хроматин и хромосомы взаимно переходят друг в друга. По химической организации как хроматин, так и хромосомы не отличаются. Химическую основу составляет дезоксирибонуклеопротеин – комплекс ДНК с белками. С помощью белков происходит многоуровневая упаковка молекул ДНК, при этом хроматин приобретает компактную форму.
Ядрышко, обычно шаровидной формы (одно или несколько), не окружено мембраной, содержит фибриллярные белковые нити и РНК. Ядрышки – не постоянные образования, они исчезают в начале деления клетки и восстанавливаются после его окончания. Ядрышки имеются только в неделящихся клетках. В ядрышках происходит формирование рибосом, синтез ядерных белков. Сами же ядрышки образуются на участках вторичных перетяжек хромосом (ядрышковых организаторах).
Ядро – обязательная часть эукариотической клетки. Диаметр ядра колеблется от 5 до 20 мкм. Главная функция ядра – хранение генетического материала в форме ДНК и передача ее дочерним клеткам при клеточном делении. Кроме того, ядро управляет белковыми синтезами, контролирует все процессы жизнедеятельности клетки. (в растительной клетке ядро описал Р.Броун в 1831г., в животной – Т.Шванн в 1838г.).
Химический состав ядра представлен, главным образом, нуклеиновыми кислотами и белками.
Строение и функции митохондрий.
Митохондрии или хондриосомы - «силовые» станции клетки, в них локализована большая часть реакций дыхания (аэробная фаза). В митохондриях происходит аккумуляция энергии дыхания в аденозинтрифосфате (АТФ). Энергия, запасаемая в АТФ, служит основным источником для физиологической деятельности клетки. Митохондрии обычно имеют удлиненную палочковидную форму длиной 4-7 мкм и диаметром 0,5-2 мкм. Число митохондрий в клетке может быть различным от 500 до 1000 и зависит от роли данного органа в процессах энергетического обмена.
Химический состав митохондрий несколько колеблется. В основном это белково-липидные органеллы. Содержание белка в них составляет 60-65%, причем структурные и ферментативные белки содержатся примерно в равной пропорции, а также около 30% липидов. Очень важно, что митохондрии содержат нуклеиновые кислоты: РНК - 1 % и ДНК -0,5%. В митохондриях имеется не только ДНК, но и вся система синтеза белка, в том числе рибосомы.
Митохондрии окружены двойной мембраной. Толщина мембран составляет 6-10 нм. Мембраны митохондрий на 70% состоят из белка. Фосфолипиды мембран представлены фосфатидтилхолином, фосфатидилэтаноламином, а также специфическими фосфолипидами, например, кардиолипином. Мембраны митохондрий не пропускают Н+ и служат барьером для их транспорта.
Между мембранами находится заполненное жидкостью перимитохондриальное пространство. Внутреннее пространство митохондрий заполняет матрикс в виде студнеобразной полужидкой массы. В матриксе сосредоточены ферменты цикла Кребса. Внутренняя мембрана дает выросты - кристы в виде пластин и трубочек, они разделяют внутреннее пространство митохондрий на отдельные отсеки. Во внутренней мембране локализована дыхательная цепь (цепь переноса электронов).
Строение и функции митохондрий представляют собой довольно сложный вопрос. Наличие органеллы характерно почти для всех ядерных организмов – как для автотрофов (растений, способных к фотосинтезу), так и для гетеротрофов, которыми являются почти все животные, некоторые растения и грибы.
Главное предназначение митохондрий – окисление органических веществ и последующее использование освободившейся в результате этого процесса энергии. По этой причине органеллы имеют также и второе (неофициальное) название – энергетические станции клетки. Иногда их называют «пластидами катаболизма».
Что такое митохондрии
Термин имеет греческое происхождение. В переводе это слово означает «нить» (mitos), «зернышко» (chondrion). Митохондрии являются постоянными органоидами, которые имеют огромное значение для нормального функционирования клеток и делают возможным существование всего организма в целом.

«Станции» имеют специфическую внутреннюю структуру, которая изменяется в зависимости от функционального состояния митохондрии. Их форма может быть двух видов – овальная или продолговатая. Последняя нередко имеет ветвящийся вид. Число органоидов в одной клетке колеблется от 150 до 1500.
Особый случай – половые клетки. В сперматозоидах присутствует всего лишь одна спиральная органелла, в то время как женских гаметах содержится в сотни тысяч больше митохондрий. В клетке органоиды не зафиксированы в одном месте, а могут передвигаться по цитоплазме, совмещаться друг с другом. Их размер составляет 0,5 мкм, длина может достигать 60 мкм, в то время как минимальный показатель – 7 мкм.

Определить размер одной «энергетической станции» – непростая задача. Дело в том, что при рассмотрении в электронный микроскоп на срез попадает только часть органеллы. Случается так, что спиральная митохондрия имеет несколько сечений, которые можно принять за отдельные, самостоятельные структуры.
Только объемное изображение позволит выяснить точное клеточное строение и понять, идет речь о 2-5 отдельных органоидах или же об одной, имеющей сложную форму митохондрии.
Особенности строения
Оболочка митохондрии состоит из двух слоев: наружного и внутреннего. Последний включает в себя различные выросты и складки, которые имеют листовидную и трубчатую форму.
Каждая мембрана имеет особенный химический состав, определенное количество тех или иных ферментов и конкретное предназначение. Наружную оболочку от внутренней отделяет межмембранное пространство толщиной 10-20 нм.
Весьма наглядно выглядит строение органеллы на рисунке с подписями.

Схема строения митохондрии
Посмотрев на схему строения, можно сделать следующее описание. Вязкое пространство внутри митохондрии называется матриксом. Его состав создает благоприятную среду для протекания в ней необходимых химических процессов. В его составе присутствуют микроскопические гранулы, которые содействуют реакциям и биохимическим процессам (например, накапливают ионы гликогена и других веществ).
В матриксе находятся ДНК, коферменты, рибосомы , т-РНК, неорганические ионы. На поверхности внутреннего слоя оболочки располагаются АТФ-синтаза и цитохромы. Ферменты способствуют таким процессам, как цикл Кребса (ЦТК), окислительное фосфорилирование и т. д.
Таким образом, главная задача органоида выполняется как матриксом, так и внутренней стороной оболочки.
Функции митохондрий
Предназначение «энергетических станций» можно охарактеризовать двумя основными задачами:
- выработка энергии: в них осуществляются окислительные процессы с последующим выделением молекул АТФ;
- хранение генетической информации;
- участие в синтезе гормонов, аминокислот и других структур.
Процесс окисления и выработки энергии проходят в несколько стадий:


Схематичный рисунок синтеза АТФ
Стоит отметить: в результате цикла Кребса (цикл лимонной кислоты) не образуются молекулы АТФ, происходит окисление молекул и выделение углекислого газа. Это промежуточный этап между гликолизом и электронтранспортной цепью.
Таблица «Функции и строение митохондрий»

От чего зависит число митохондрий в клетке
Превалирующее число органоидов скапливается рядом с теми участками клетки, где возникает необходимость в энергетических ресурсах. В частности, большое количество органелл собирается в зоне нахождения миофибрилл, которые являются частью мышечных клеток, обеспечивающих их сокращение.

В мужских половых клетках структуры локализуются вокруг оси жгутика – предполагается, что потребность в АТФ обусловлена постоянным движением хвоста гаметы. Точно так же выглядит расположение митохондрий у простейших, которые для передвижения используют специальные реснички – органеллы скапливаются под мембраной у их основания.
Что касается нервных клеток, то локализация митохондрий наблюдается вблизи синапсов, через которые передаются сигналы нервной системы. В клетках, синтезирующих белки, органеллы скапливаются в зонах эргастоплазмы – они поставляют энергию, которая обеспечивает данный процесс.
Кто открыл митохондрии
Свое название клеточная структура обрела в 1897-1898 годах благодаря К. Бренду. Связь процессов клеточного дыхания с митохондриями сумел доказать Отто Вагбург в 1920 году.
Заключение
Митохондрии являются важнейшей составляющей живой клетки, выступая в роли энергетической станции, которая производит молекулы АТФ, обеспечивая тем самым процессы клеточной жизнедеятельности.
Работа митохондрий основана на окислении органических соединений, в результате чего происходит генерация энергетического потенциала.
Основной функцией митохондрий является синтез АТФ - универсальной формы химической энергии в любой живой клетке. Как и у прокариот, данная молекула может образовываться двумя путями: в результате субстратного фосфорилирования в жидкой фазе (например, при гликолизе) или в процессе мембранного фосфорилирования, связанного с использованием энергии трансмембранного электрохимического градиента (англ.)русск. протонов (ионов водорода). Митохондрии реализуют оба эти пути, первый из которых характерен для начальных процессов окисления субстрата и происходит в матриксе, а второй завершает процессы энергообразования и связан с кристами митохондрий. При этом своеобразие митохондрий как энергообразующих органелл эукариотической клетки определяет именно второй путь генерации АТФ, получивший название «хемиосмотического сопряжения». По сути это последовательное превращение химической энергии восстанавливающих эквивалентов НАДН в электрохимический протонный градиент ΔμН + по обе стороны внутренней мембраны митохондрии, что приводит в действие мембранно-связанную АТФ-синтетазу и завершается образованием макроэргической связи в молекуле АТФ.
В целом весь процесс энергообразования в митохондриях может быть разбит на четыре основные стадии, первые две из которых протекают в матриксе, а две последние - на кристах митохондрий:
- Превращение поступивших из цитоплазмы в митохондрию пирувата и жирных кислот в ацетил-СоА;
- Окисление ацетил-СоА в цикле Кребса, ведущее к образованию НАДН;
- Перенос электронов с НАДН на кислород по дыхательной цепи;
- Образование АТФ в результате деятельности мембранного АТФ-синтетазного комплекса.
Ещё в цитоплазме в серии из 10 отдельных ферментативных реакций шестиуглеродная молекула глюкозы частично окисляется до двух трёхуглеродных молекул пирувата с образованием двух молекул АТФ. Затем пируват переносится из цитозоля через наружную и внутреннюю мембраны в матрикс, где первоначально превращается в ацетил-СоА. Этот процесс катализируется крупным пируватдегидрогеназным комплексом, имеющим размер, сопоставимый с размером рибосомы, и состоящим из трёх ферментов, пяти коферментов и двух регуляторных белков. Точно также жирные кислоты, полученные при расщеплении нерастворимых триглицеридов в цитоплазме, переносятся в митохондриальный матрикс в виде ацетил-СоА-производных.
На следующем этапе, также протекающем в матриксе митохондрии, ацетил-СоА полностью окисляется в цикле Кребса. В его работе задействованы четыре отдельных фермента, за каждый цикл обеспечивающие укорочение углеводородной цепи на два атома углерода, которые в дальнейшем превращаются в СО 2 . Этот процесс обеспечивает образование одной молекулы АТФ, а также НАДН - высокоэнергетического промежуточного соединения, которое легко отдаёт электроны в цепь переноса электронов на кристах митохондрий.
Дальнейшие процессы энергообразования в митохондрии происходят на её кристах и связаны с переносом электронов от НАДН к кислороду. В соответствии с тем, что потребление кислорода в качестве окислителя обычно называют «внутриклеточным дыханием», электронно-транспортную цепь ферментов, осуществляющих последовательный перенос электронов от НАДН к кислороду, часто называют «дыхательной цепью». При этом трансформация энергии окисления осуществляется ферментами, расположенными на кристах митохондрий и осуществляющими векторный (направленный по отношению к сторонам мембраны) перенос протонов водорода из матрикса митохондрии в межмембранное пространство. В этом состоит принципиальное отличие работы оксидоредуктаз дыхательной цепи от функционирования ферментов, катализирующих реакции в гомогенном (изотропном) растворе, где вопрос о направлении реакции в пространстве не имеет смысла.
Весь процесс переноса электрона по дыхательной цепи может быть разбит на три стадии, каждая из которых катализируется отдельным трансмембранным липопротеидным комплексом (I, III и IV), встроенным в мембрану кристы митохондрии. В состав каждого из названных комплексов входят следующие компоненты:
- Большой олигомерный фермент, катализирующий перенос электронов;
- Небелковые органические (простетические) группы, принимающие и высвобождающие электроны;
- Белки, обеспечивающие движение электронов.
Каждый из этих комплексов осуществляет перенос электронов от донора к акцептору по градиенту редокс-потенциала через ряд последовательно функционирующих переносчиков. В качестве последних в дыхательной цепи митохондрий функционируют мигрирующие в плоскости мембраны жирорастворимые молекулы убихинона, а также небольшие (молекулярная масса 13кДа) водорастворимые белки, содержащие ковалентно связанный гем и называемые «цитохромами с ». При этом три из пяти компонентов, составляющих дыхательную цепь, работают так, что перенос электронов сопровождается переносом протонов через мембрану крист митохондрий в направлении из матрикса в межмембранное пространство.
Дыхательная цепь начинается с комплекса I (НАДН-убихинон-оксидоредуктаза), состоящего из 16-26 полипептидных цепей и имеющего молекулярную массу около 850 кДа. Функциональная активность этого комплекса определяется тем, что он содержит в своём составе более 20 атомов железа, упакованных в ячейки из атомов серы, а также флавин (Фл - производное витамина рибофлавина). Комплекс I катализирует окисление НАДН, отщепляя от него два электрона, которые после «путешествия» по окислительно-восстановительным компонентам комплекса I попадают на молекулу-переносчик, в качестве которой выступает убихинон (Q). Последний способен ступенчато восстанавливаться, принимая на себя по два электрона и протона и, таким образом, превращаясь в восстановленную форму - убихинол (QH 2).
Энергетический потенциал (запас энергии) в молекуле убихинола существенно ниже, чем в молекуле НАДН, а разница в подобной энергии временно запасается в виде особого вида - электрохимического протонного градиента. Последний возникает в результате того, что перенос электронов по простетическим группам комплекса I, ведущий к снижению энергетического потенциала электронов, сопровождается трансмембранным переносом двух протонов из матрикса в межмембранное пространство митохондрии.
Восстановленный убихинол мигрирует в плоскости мембраны, где достигает второго фермента дыхательной цепи - комплекса III (bc 1 ). Последний представляет собой димер из субъединиц b и c 1 с молекулярной массой более 300 кДа, сформированный из восьми полипептидных цепей и содержащий атомы железа как в серных ячейках, так и в виде комплексов с гемами b (I), b (II) и c 1 - сложными гетероциклическими молекулами с четырьмя атомами азота, расположенными по углам металлосвязывающего квадрата. Комплекс III катализирует реакцию восстановления убихинола до убихинона с передачей электронов на атом железа второй молекулы переносчика (находящегося в межмембранном пространстве цитохрома c ). Отщепляющиеся при этом от убихинола два протона водорода освобождаются в межмембранное пространство, продолжая формирование электрохимического градиента. Наконец, ещё два протона водорода переносятся в межмембранное пространство митохондрии за счёт энергии электронов, проходящих по простетических группам комплекса III.
Последняя стадия катализируется комплексом IV (цитохром c -оксидаза) с молекулярной массой около 200 кДа, состоящим из 10-13 полипептидных цепей и, помимо двух различных гемов, включающим также несколько атомов меди, прочно связанных с белками. При этом электроны, отбираемые у восстановленного цитохрома c , пройдя по атомам железа и меди в составе комплекса IV, попадают на связанный в активном центре этого фермента кислород, что приводит к образованию воды.
Таким образом, суммарная реакция, катализируемая ферментами дыхательной цепи, состоит в окислении НАДН кислородом с образованием воды. По сути этот процесс заключается в ступенчатом переносе электронов между атомами металлов, присутствующих в простетических группах белковых комплексов дыхательной цепи, где каждый последующий комплекс обладает более высоким сродством к электрону, чем предыдущий. При этом сами электроны передаются по цепи до тех пор, пока не соединятся с молекулярным кислородом, обладающим наибольшим сродством к электронам. Освобождаемая же при этом энергия запасается в виде электрохимического (протонного) градиента по обе стороны внутренней мембраны митохондрий. При этом считается, что в процессе транспорта по дыхательной цепи пары электронов перекачивается от трёх до шести протонов.
Завершающим этапом функционирования митохондрии является генерация АТФ, осуществляемая встроенным во внутреннюю мембрану специальным макромолекулярным комплексом с молекулярной массой 500 кДа. Этот комплекс, называемый АТФ-синтетазой, как раз и катализирует синтез АТФ путём конверсии энергии трансмембранного электрохимического градиента протонов водорода в энергию макроэргической связи молекулы АТФ.
АТФ-синтаза
В структурно-функциональном плане АТФ-синтаза состоит из двух крупных фрагментов, обозначаемых символами F 1 и F 0 . Первый из них (фактор сопряжения F 1) обращён в сторону матрикса митохондрии и заметно выступает из мембраны в виде сферического образования высотой 8 нм и шириной 10 нм. Он состоит из девяти субъединиц, представленных пятью типами белков. Полипептидные цепи трёх субъединиц α и стольких же субъединиц β уложены в похожие по строению белковые глобулы, которые вместе образуют гексамер (αβ) 3 , имеющий вид слегка приплюснутого шара. Подобно плотно уложенным долькам апельсина, последовательно расположенные субъединицы α и β образуют структуру, характеризующуюся осью симметрии третьего порядка с углом поворота 120°. В центре этого гексамера находится субъединица γ, которая образована двумя протяжёнными полипептидными цепями и напоминает слегка деформированный изогнутый стержень длиной около 9 нм. При этом нижняя часть субъединицы γ выступает из шара на 3 нм в сторону мембранного комплекса F 0 . Также внутри гексамера находится минорная субъединица ε, связанная с γ. Последняя (девятая) субъединица обозначается символом δ и расположена на внешней стороне F 1 .
Мембранная часть АТФ-синтазы, называемая фактором сопряжения F 0 , представляет собой гидрофобный белковый комплекс, пронизывающий мембрану насквозь и имеющий внутри себя два полуканала для прохождения протонов водорода. Всего в состав комплекса F 0 входит одна белковая субъединица типа а , две копии субъединицы b , а также от 9 до 12 копий мелкой субъединицы c . Субъединица а (молекулярная масса 20 кДа) полностью погружена в мембрану, где образует шесть пересекающих её α-спиральных участков. Субъединица b (молекулярная масса 30 кДа) содержит лишь один сравнительно короткий погружённый в мембрану α-спиральный участок, а остальная её часть заметно выступает из мембраны в сторону F 1 и закрепляется за расположенную на её поверхности субъединицу δ. Каждая из 9-12 копий субъединицы c (молекулярная масса 6-11 кДа) представляет собой сравнительно небольшой белок из двух гидрофобных α-спиралей, соединённых друг с другом короткой гидрофильной петлёй, ориентированной в сторону F 1 , а все вместе образуют единый ансамбль, имеющий форму погружённого в мембрану цилиндра. Выступающая из комплекса F 1 в сторону F 0 субъединица γ как раз и погружена внутрь этого цилиндра и достаточно прочно зацеплена за него.
Таким образом, в молекуле АТФ-синтазы можно выделить две группы белковых субъединиц, которые могут быть уподоблены двум деталям мотора: ротору и статору. «Статор» неподвижен относительно мембраны и включает в себя шарообразный гексамер (αβ) 3 , находящуюся на его поверхности и субъединицу δ, а также субъединицы a и b мембранного комплекса F 0 . Подвижный относительно этой конструкции «ротор» состоит из субъединиц γ и ε, которые, заметно выступая из комплекса (αβ) 3 , соединяются с погружённым в мембрану кольцом из субъединиц c .
Способность синтезировать АТФ - свойство единого комплекса F 0 F 1 , сопряжённого с переносом протонов водорода через F 0 к F 1 , в последнем из которых как раз и расположены каталитические центры, осуществляющие преобразование АДФ и фосфата в молекулу АТФ. Движущей же силой для работы АТФ-синтазы является протонный потенциал, создаваемый на внутренней мембране митохондрий в результате работы цепи электронного транспорта.
Сила, приводящая в движение «ротор» АТФ-синтазы, возникает при достижении разности потенциалов между наружной и внутренней сторонами мембраны > 220 мВ и обеспечивается потоком протонов, протекающих через специальный канал в F 0 , расположенный на границе между субъединицами a и c . При этом путь переноса протонов включает в себя следующие структурные элементы:
- Два расположенных несоосно «полуканала», первый из которых обеспечивает поступление протонов из межмембранного пространства к существенно важным функциональным группам F 0 , а другой обеспечивает их выход в матрикс митохондрии;
- Кольцо из субъединиц c , каждая из которых в своей центральной части содержит протонируемую карбоксильную группу, способную присоединять H + из межмембранного пространства и отдавать их через соответствующие протонные каналы. В результате периодических смещений субъединиц с , обусловленных потоком протонов через протонный канал происходит поворот субъединицы γ, погружённой в кольцо из субъединиц с .
Таким образом, каталитическая активность АТФ-синтазы непосредственно связана с вращением её «ротора», при котором поворот субъединицы γ вызывает одновременное изменение конформации всех трёх каталитических субъединиц β, что в конечном счёте и обеспечивает работу фермента. При этом в случае образования АТФ «ротор» крутится по часовой стрелке со скоростью четыре оборота в секунду, а само подобное вращение происходит дискретными скачками по 120°, каждый из которых сопровождается образованием одной молекулы АТФ.
Непосредственная функция синтеза АТФ локализована на β-субъединицах сопрягающего комплекса F 1 . При этом самым первым актом в цепи событий, приводящих к образованию АТФ, является связывание АДФ и фосфата с активным центром свободной β-субъединицы, находящейся в состоянии 1. За счёт энергии внешнего источника (тока протонов) в комплексе F 1 происходят конформационные изменения, в результате которых АДФ и фосфат становятся прочно связанными с каталитическим центром (состояние 2), где становится возможным образование ковалентной связи между ними, ведущей к образованию АТФ. На данной стадии АТФ-синтазы ферменту практически не требуется энергии, которая будет необходима на следующем этапе для освобождения прочно связанной молекулы АТФ из ферментативного центра. Поэтому следующий этап работы фермента заключается в том, чтобы в результате энергозависимого структурного изменения комплекса F 1 каталитическая β-субъединица, содержащая прочно связанную молекулу АТФ, перешла в состояние 3, в котором связь АТФ с каталитическим центром ослаблена. В результате этого молекула АТФ покидает фермент, а β-субъединица возвращается в исходное состояние 1, благодаря чему обеспечивается цикличность работы фермента.
Работа АТФ-синтазы связана с механическими движениями её отдельных частей, что позволило отнести этот процесс к особому типу явлений, названных «вращательным катализом». Подобно тому, как электрический ток в обмотке электродвигателя приводит в движение ротор относительно статора, направленный перенос протонов через АТФ-синтетазу вызывает вращение отдельных субъединиц фактора сопряжения F 1 относительно других субъединиц ферментного комплекса, в результате чего это уникальное энергообразующее устройство совершает химическую работу - синтезирует молекулы АТФ. В дальнейшем АТФ поступает в цитоплазму клетки, где расходуется на самые разнообразные энергозависимые процессы. Подобный перенос осуществляется специальным встроенным в мембрану митохондрий ферментом АТФ/АДФ-транслоказой, который обменивает вновь синтезированную АТФ на цитоплазматическую АДФ, что гарантирует сохранность фонда адениловых нуклеотидов внутри митохондрий.
Митохондрии растительной клетки. Их структура и функции
Форма − округлые или гантелевидные тельца.
Размеры − длина 1-5 мкм, диаметром 0,4-0,5 мкм.
Количество в клетке − от десятков до 5 000.
Структура . Состоят в основном из белка (60-65 %) и липидов (30 %). Это двухмембранные органоиды. Толщина наружной и внутренней мембран − 5-6 нм каждая. Перимитохондриальное пространство (промежуток между мемранами) заполнено жидкостью типа сыворотки. Внутренняя мембрана образует различной формы складки − кристы . На внутренней поверхности внутренней мембраны расположены грибовидные частицы − оксисомы, содержащие окислительные ферменты. Внутреннее содержимое митохондрий − матрикс . В матриксе содержатся рибосомы и митохондриальная ДНК (0,5 %), которая имеет кольцевое строение и отвечает за синтез белков митохондрий. Митохондрии имеют все типы РНК (1 %), делятся независимо от деления ядра, в клетке образуются от предсуществующих митохондрий путем деления или почкования. Полупериод жизни митохондрий − 5−10 дней.
Функции . Митохондрии являются центрами энергетической активности клеток. В митохондриях функционируют системы аэробного дыхания и окислительного фосфорелирования. Во внутренней мембране митохондрий локализованы компоненты электронтранспортной цепи и АТФ-синтетазные комплексы, осуществляющие транспорт электронов и протонов и синтез АТФ. В матриксе располагаются системы окисления ди- и трикарбоновых кислот, ряд систем синтеза липидов, аминокислот и др.
Митохондрии способны передвигаться к местам усиленного потребления энергии. Они могут ассоциировать друг с другом путем тесного сближения или при помощи тяжей. При анаэробном дыхании митохондрии исчезают.
Митохондрии имеют округлую и продолговатую форму диаметром 0,4–0,5 мкм и длиной 1–5 мкм (рис. 1.3).
Количество митохондрий варьирует от единиц до 1 500–2 000 на растительную клетку.
Митохондрии ограничены двумя мембранами: наружной и внутренней, толщина каждой из них 5–6 нм. Наружная мембрана выглядит растянутой, а внутренняя образует складки, называемые гребнями (кристами), различной формы. Пространство между мембранами, в состав которого входит также внутреннее пространство крист, называется межмембранным (перимитохондриальным) пространством. Оно служит средой для внутренней мембраны и матрикса митохондрий.
Митохондрии в целом содержат 65–70 % белка, 25–30 % липидов и небольшое количество нуклеиновых кислот. 70 % от общего содержания липидов составляют фосфолипиды (фосфатидилхолин и фосфатидилэтаноламин). Жирнокислотный состав характеризуется высоким содержанием насыщенных жирных кислот, обеспечивающих «жесткость» мембраны.
В митохондриях локализованы системы аэробного дыхания и окислительного фосфорилирования. В результате дыхания расщепляются органические молекулы, и высвобождается энергия с передачей ее на молекулу АТФ.
Митохондрии содержат белки, РНК, тяжи ДНК, рибосомы, сходные с бактериальными, и различные растворенные вещества. ДНК существует в виде кольцевых молекул, располагающихся в одном или нескольких нуклеотидах.
Пластиды, наряду с вакуолями и клеточной оболочкой – характерные компоненты растительных клеток. Каждая пластида окружена собственной оболочкой, состоящей из двух элементарных мембран. Внутри пластид различают мембранную систему и более или менее гомогенное вещество – строму. Внутренняя структура хлоропласта довольно сложна. Строма пронизана развитой системой мембран, имеющих форму плоских пузырьков, называемых тилакоидами.Тилакоиды собраны в стопки – граны, напоминающие столбики монет.
 |
Хлоропласты, в которых протекает фотосинтез, содержат хлорофиллы и каротиноиды. Размер – 4–5 мкм. В одной клетке мезофилла листа может содержаться 40–50 хлоропластов, в мм 2 листа – около 500 000. В цитоплазме хлоропласты обычно располагаются параллельно клеточной оболочке.
Хлорофиллы и каротиноиды встроены в тилакоидные мембраны. Хлоропласты зеленых растений и водорослей часто содержат зерна крахмала и мелкие липидные (жировые) капли. Крахмальные зерна – это временные хранилища продуктов фотосинтеза. Они могут исчезнуть из хлоропластов, находящихся в темноте всего лишь 24 ч и появиться вновь уже через 3–4 ч после переноса растений на свет.
В изолированных хлоропластах осуществляется синтез РНК, который обычно контролируется только хромосомной ДНК. Образование хлоропластов и синтез находящихся в них пигментов в значительной степени контролируется хромосомной ДНК, малопонятным образом взаимодействующей с ДНК хлоропластов. Тем не менее, в отсутствие собственной ДНК хлоропласты не формируются.
Хлоропласты могут считаться основными клеточными органеллами, поскольку первыми стоят в цепи преобразования солнечной энергии, в результате которого мы получаем пищу и топливо. В хлоропластах протекают не только фотосинтез. Они участвуют в синтезе аминокислот и жирных кислот, служат хранилищем временных запасов крахмала.
Хромопласты (от греческого сhroma – цвет) – пигментированные пластиды. Многообразные по форме хромопласты не содержат хлорофилла, но синтезируют и накапливают каротиноиды, которые придают желтую, оранжевую и другую окраску. Корнеплоды моркови, плоды томатов окрашены пигментами, которые находятся в хромопластах.
Лейкопласты являются местом накопления запасного вещества – крахмала. Особенно много лейкопластов в клетках клубней картофеля. На свету лейкопласты могут преобразовываться в хлоропласты (клубни картофеля зеленеют). Осенью хлоропласты преобразуются в хромопласты и зеленые листья, и плоды желтеют и краснеют.
Характерные для подавляющего большинства клеток. Основная функция – это окисление органических соединений и производство АТФ-молекул из освободившейся энергии. Маленькая митохондрия является главной энергетической станцией всего организма.
Происхождение митохондрий
Среди ученых на сегодняшний день весьма популярно мнение, что митохондрия появилась в клетке не самостоятельно в ходе эволюции. Скорее всего, это произошло из-за захвата примитивной клеткой, не способной на тот момент самостоятельно использовать кислород, бактерии, которая умела это и, соответственно, была превосходным источником энергии. Подобный симбиоз оказался успешным и закрепился в последующих поколениях. В пользу этой теории указывает наличие в митохондриях собственной ДНК.
Как устроены митохондрии
Митохондрия обладает двумя мембранами: наружной и внутренней. Главная функция наружной мембраны – это отделение органоида от цитоплазмы клетки. Она состоит из билипидного слоя и белков, пронизывающих его, через которые и осуществляется транспорт молекул и ионов, необходимых для работы. В то время как гладкая, внутренняя образует многочисленные складки – кристы, которые существенно увеличивают ее площадь. Внутренняя мембрана по большей части состоит из белков, среди которых присутствуют ферменты дыхательной цепи, транспортные белки и крупные АТФ-синтетазные комплексы. Именно в этом месте происходит синтез АТФ. Между наружной и внутренней мембраной находится межмембранное пространство с присущими ему ферментами.
Внутреннее пространство митохондрий называется матрикс. Здесь расположены ферментные системы окисления жирных кислот и пирувата, ферменты цикла Кребса, а также наследственный материал митохондрий – ДНК, РНК и белоксинтезирующий аппарат.
Для чего нужны митохондрии
Главной функцией митохондрий является синтез универсальной формы химической энергии – АТФ. Также они принимают участие в цикле трикарбоновых кислот, превращая пируват и жирные кислоты в ацетил-CoA, а затем окисляя его. В этом органоиде хранится и передается по наследству митохондриальная ДНК, кодирующая воспроизводство тРНК, рРНК и некоторых белков, необходимых для нормального функционирования митохондрий.