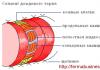
Механизм действия тиреоидных гормонов на клетки-мишени
Гормоны T3 и Т4 являются жирорастворимыми гормонами, которые транспортируются через мембрану в цитоплазму клетки-мишени (шаг 1) и связываются с тиреоидными рецепторами в ядре (шаг 2). Образованный комплекс ГР взаимодействует с ДНК (шаг 3), стимулирует процессы транскрипции - образование мРНК (шаг 4) и как следствие - синтез новых белков на рибосомах (шаг 5), что приводит к изменению функции клетки-мишени (шаг 6) (рис. 6.13).
Роль гормонов щитовидной железы в процессах роста, психического развития и метаболизма
Влияние гормонов на рост. Тиреоидные гормоны, как синергисты гормона роста и соматомединов (ИФР-I), в физиологических концентрациях стимулируют рост и развитие скелета, путем потенциации синтеза белков в клетках-мишенях, в том числе в хондроцитах, скелетных мышцах.
Гормоны также способствуют оссификации костей - закрыванию эпифизарных ростовых зон. При их недостатке ростовые зоны долго не закрываются и развитие костей отстает от хронологического возраста.
Влияние гормонов на ЦНС. Развитие ЦНС у детей после рождения осуществляется с обязательным учас-
РИС. 6.13. Схема механизма действия тиреоидных гормонов и их основные влияния на функции организма. 1-6 - последовательность реакции гормона со структурами ядра и системой синтеза новых белков
тю тиреоидных гормонов. Они способствуют миелинизации и разветвления отростков нейронов головного мозга, развития психических функций. Наибольшее влияние проявляется на кору большого мозга, базальные ганглии, завиток. При отсутствии тиреоидных гормонов в перинатальном периоде возникает умственная отсталость - кретинизм. Существует очень короткий период времени после рождения, когда заместительная терапия гормонами может способствовать нормальному психическому развитию. Поэтому важно выявлять дефицит гормонов еще до рождения ребенка.
У взрослых людей нормальные психические функции, память, скорость рефлекторных реакций поддерживаются с участием тиреоидных гормонов непосредственно и косвенно - благодаря увеличению количества адренорецепторов в нейронах ЦНС.
Люди, у которых наблюдается избыток тиреоидных гормонов, становятся раздражительными, неугомонными, скорость мыслительных процессов ускоряется. У людей с недостатком тиреоидных процессов мыслительные процессы замедляются, память ухудшается, скорость рефлекторных реакций уменьшается.
Влияние гормонов на интенсивность метаболизма. Интенсивность метаболизма в состоянии покоя под влиянием гормонов возрастает, особенно это заметно в условиях избытка тиреоидных гормонов. Увеличение интенсивности метаболизма происходит почти во всех клетках-мишенях, за исключением головного мозга, яичек, лимфатических узлов, селезенки, аденогипофиза. Повышается поглощение кислорода, теплообразования.
Рост интенсивности метаболизма под действием тиреоидных гормонов может в своей основе иметь их влияние на синтез клеточного ферментного белка - натрий калиевой АТФ-азы, расположенной в мембранах клеток. В свою очередь, интенсивная работа натрий-калиевых насосов увеличивает интенсивность метаболизма.
Влияние гормонов на углеводный метаболизм. Тиреоидные гормоны в физиологических концентрациях потенцируют действие инсулина и способствуют гликогенеза и утилизации глюкозы.
При увеличении концентрации гормонов (во время стресса или фармакологическим путем) развивается гипергликемия за счет потенциации гликогенолиза, вызванного адреналином. Растет глюконеогенез, окисления глюкозы и ее всасывания в кишечнике вторичным активным транспортом.
Влияние гормонов на белковый метаболизм. Тиреоидные гормоны в физиологических концентрациях имеют анаболическим действием - стимулируют синтез белков, однако в больших концентрациях вызывают их катаболизм.
Влияние гормонов на жировой метаболизм. Тиреоидные гормоны стимулируют все аспекты жирового метаболизма - синтез липидов, их мобилизацию и использование. Увеличение их концентрации приводит к липолиза - уменьшение в крови концентрации триглицеридов, фосфолипидов и рост свободных жирных кислот и глицерина. Под влиянием гормонов повышается количество рецепторов липопротеинов низкой плотности (ЛПНП) и падает число холестероловых - в печени. Это приводит к увеличению выделения холестерина из организма, уменьшение его уровня в крови.
Метаболизм жирорастворимых витаминов также находится под влиянием тиреоидных гормонов - они необходимы для синтеза витамина А из каротина и превращение его в ретинен.
Влияние гормонов на автономную нервную систему заключается в том, что в клетках-мишенях увеличивается количество бета-адренорецепторов, которые синтезируются под влиянием тиреоидных гормонов, что приводит к усилению эффекта катехоламинов в эффекторных клетках.
Влияние гормонов на висцеральные системы. Система кровообращения. Частота сокращения сердца ускоряется благодаря увеличению количества β-адренорецепторов в пейсмекера и усиление влияния катехоламинов; сила сокращения - повышается в результате увеличения в кардиомиоцитах пула тяжелых цепей α-миозина, имеющих высокую активность АТФ-азы.
Система дыхания. Вентиляция легких углубляется, что является приспособительной реакцией к росту поглощения кислорода при увеличении интенсивности метаболизма.
Организм человека существует как единое целое благодаря системе внутренних связей, которая обеспечивает передачу информации от одной клетки к другой в одной и той же ткани или между разными тканями. Без этой системы невозможно поддерживать гомеостаз. В передаче информации между клетками в многоклеточных живых организмах, принимают участие три системы: ЦЕНТРАЛЬНАЯ НЕРВНАЯ СИСТЕМА (ЦНС), ЭНДОКРИННАЯ СИСТЕМА (ЖЕЛЕЗЫ ВНУТРЕННЕЙ СЕКРЕЦИИ) и ИММУННАЯ СИСТЕМА.
Способы передачи информации во всех названных системах - химические. Посредниками при передаче информации могут быть СИГНАЛЬНЫЕ молекулы.
К таким сигнальным молекулам относятся четыре группы веществ: ЭНДОГЕННЫЕ БИОЛОГИЧЕСКИ АКТИВНЫЕ ВЕЩЕСТВА (медиаторы иммунного ответа, факторы роста и др.), НЕЙРОМЕДИАТОРЫ, АНТИТЕЛА (иммуноглобулины) и ГОРМОНЫ.
Б И О Х И М И Я Г О Р М О Н О В
ГОРМОНЫ - это биологически активные вещества, которые синтезируются в малых количествах в специализированнных клетках эндокринной системы и через циркулирующие жидкости (например, кровь) доставляются к клеткам-мишеням, где оказывают свое регулирующее действие.
Гормоны, как и другие сигнальные молекулы, обладают некоторыми общими свойствами.
ОБЩИЕ СВОЙСТВА ГОРМОНОВ.
1) выделяются из вырабатывающих их клеток во внеклеточное пространство;
2) не являются структурными компонентами клеток и не используются как источник энергии.
3) способны специфически взаимодействовать с клетками, имеющими рецепторы для данного гормона.
4) обладают очень высокой биологической активностью - эффективно действуют на клетки в очень низких концентрациях (около 10 -6 - 10 -11 моль/л).
МЕХАНИЗМЫ ДЕЙСТВИЯ ГОРМОНОВ.
Гормоны оказывают влияние на клетки-мишени.
КЛЕТКИ-МИШЕНИ - это клетки, которые специфически взаимодействуют с гормонами с помощью специальных белков-рецепторов. Эти белки-рецепторы располагаются на наружной мембране клетки, или в цитоплазме, или на ядерной мембране и на других органеллах клетки.
БИОХИМИЧЕСКИЕ МЕХАНИЗМЫ ПЕРЕДАЧИ СИГНАЛА ОТ ГОРМОНА В КЛЕТКУ-МИШЕНЬ.
Любой белок-рецептор состоит, минимум из двух доменов (участков), которые обеспечивают выполнение двух функций:
- "узнавание" гормона;
Преобразование и передачу полученного сигнала в клетку.
Каким образом белок-рецептор узнает ту молекулу гормона, с которой он может взаимодействовать?
Один из доменов белка-рецептора имеет в своем составе участок, комплементарный какой-то части сигнальной молекулы. Процесс связывания рецептора с сигнальной молекулой похож на процесс образования фермент-субстратного комплекса и может определяется величиной константы сродства.
Большинство рецепторов изучены недостаточно, потому что их выделение и очистка очень сложные, а содержание каждого вида рецепторов в клетках очень низкое. Но известно, что гормоны взаимодействуют со своими рецепторами физико-химическим путем. Между молекулой гормона и рецептором формируются электростатические и гидрофобные взаимодействия. При связывании рецептора с гормоном происходят конформационные изменения белка-рецептора и комплекс сигнальной молекулы с белком-рецептором активируется. В активном состоянии он может вызывать специфические внутриклеточные реакции в ответ на принятый сигнал. Если нарушен синтез или способность белков-рецепторов связываться с сигнальными молекулами, возникают заболевания - эндокринные нарушения. Есть три типа таких заболеваний:
1. Связанные с недостаточностью синтеза белков-рецепторов.
2. Связанные с изменением структуры рецептора - генетических дефекты.
3. Связанные с блокированием белков-рецепторов антителами.
Гормоны оказывают влияние на клетки-мишени.
Клетки-мишени — это клетки, которые специфически взаимодействуют с гормонами с помощью специальных белков-рецепторов. Эти белки-рецепторы располагаются на наружной мембране клетки, или в цитоплазме, или на ядерной мембране и на других органеллах клетки.
Биохимические механизмы передачи сигнала от гормона в клетку-мишень.
Любой белок-рецептор состоит, минимум из двух доменов (участков), которые обеспечивают выполнение двух функций:
узнавание гормона;
преобразование и передачу полученного сигнала в клетку.
Каким образом белок-рецептор узнает ту молекулу гормона, с которой он может взаимодействовать?
Один из доменов белка-рецептора имеет в своем составе участок, комплементарный какой-то части сигнальной молекулы. Процесс связывания рецептора с сигнальной молекулой похож на процесс образования фермент-субстратного комплекса и может определяется величиной константы сродства.
Большинство рецепторов изучены недостаточно, потому что их выделение и очистка очень сложные, а содержание каждого вида рецепторов в клетках очень низкое. Но известно, что гормоны взаимодействуют со своими рецепторами физико-химическим путем. Между молекулой гормона и рецептором формируются электростатические и гидрофобные взаимодействия. При связывании рецептора с гормоном происходят конформационные изменения белка-рецептора и комплекс сигнальной молекулы с белком-рецептором активируется. В активном состоянии он может вызывать специфические внутриклеточные реакции в ответ на принятый сигнал. Если нарушен синтез или способность белков-рецепторов связываться с сигнальными молекулами, возникают заболевания — эндокринные нарушения.
Есть три типа таких заболеваний.
Связанные с недостаточностью синтеза белков-рецепторов.
Связанные с изменением структуры рецептора — генетических дефекты.
Связанные с блокированием белков-рецепторов антителами.
Механизмы действия гормонов на клетки-мишени.
В зависимости от строения гормона существуют два типа взаимодействия. Если молекула гормона липофильна, (например, стероидные гормоны), то она может проникать через липидный слой наружной мембраны клеток-мишеней. Если молекула имеет большие размеры или является полярной, то ее проникновение внутрь клетки невозможно. Поэтому для липофильных гормонов рецепторы находятся внутри клеток-мишеней, а для гидрофильных — рецепторы находятся в наружной мембране.
Для получения клеточного ответа на гормональный сигнал в случае гидрофильных молекул действует внутриклеточный механизм передачи сигнала. Это происходит с участием веществ, которых называют вторыми посредниками. Молекулы гормонов очень разнообразны по форме, а "вторые посредники" — нет.
Надежность передачи сигнала обеспечивает очень высокое сродство гормона к своему белку-рецептору.
Что такое посредники, которые участвуют во внутриклеточной передаче гуморальных сигналов?
Это циклические нуклеотиды (цАМФ и цГМФ), инозитолтрифосфат, кальций-связывающий белок — кальмодулин, ионы кальция, ферменты, участвующие в синтезе циклических нуклеотидов, а также протеинкиназы — ферменты фосфорилирования белков. Все эти вещества участвуют в регуляции активности отдельных ферментных систем в клетках-мишенях.
Разберем более подробно механизмы действия гормонов и внутриклеточных посредников.
Существует два главных способа передачи сигнала в клетки-мишени от сигнальных молекул с мембранным механизмом действия:
аденилатциклазная (или гуанилатциклазная) системы;
фосфоинозитидный механизм.
Аденилатциклазная система.
Основные компоненты: мембранный белок-рецептор, G-белок, фермент аденилатциклаза, гуанозинтрифосфат, протеинкиназы.
Кроме того, для нормального функционирования аденилатциклазной системы, требуется АТФ.
Белок-рецептор, G-белок, рядом с которым располагаются ГТФ и фермент (аденилатциклаза) встроены в мембрану клетки.
До момента действия гормона эти компоненты находятся в диссоциированнном состоянии, а после образования комплекса сигнальной молекулы с белком-рецептором происходят изменения конформации G-белка. В результате одна из субъединиц G-белка приобретает способность связываться с ГТФ.
Комплекс "G-белок-ГТФ" активирует аденилатциклазу. Аденилатциклаза начинает активно превращать молекулы АТФ в ц-АМФ.
ц-АМФ обладает способностью активировать особые ферменты — протеинкиназы, которые катализируют реакции фосфорилирования различных белков с участием АТФ. При этом в состав белковых молекул включаются остатки фосфорной кислоты. Главным результатом этого процесса фосфорилирования является изменение активности фосфорилированного белка. В различных типах клеток фосфорилированию в результате активации аденилат-циклазной системы подвергаются белки с разной функциональной активностью. Например, это могут быть ферменты, ядерные белки, мембранные белки. В результате реакции фосфорилирования белки могут становятся функционально активными или неактивными.
Такие процессы будут приводить к изменениям скорости биохимических процессов в клетке-мишени.
Активация аденилатциклазной систтемы длится очень короткое время, потому что G-белок после связывания с аденилатциклазой начинает проявлять ГТФ-азную активность. После гидролиза ГТФ G-белок восстанавливает свою конформацию и перестает активировать аденилатциклазу. В результате прекращается реакция образования цАМФ.
Кроме участников аденилатциклазной системы в некоторых клетках-мишенях имеются белки-рецепторы, связанные с G-белками, которые приводят к торможению аденилатциклазы. При этом комплекс "GTP-G-белок" ингибирует аденилатциклазу.
Когда останавливается образование цАМФ, реакции фосфорилирования в клетке прекращаются не сразу: пока продолжают существовать молекулы цАМФ — будет продолжаться и процесс активации протеинкиназ. Для того, чтобы прекратить действие цАМФ, в клетках существует специальный фермент — фосфодиэстераза, который катализирует реакцию гидролиза 3′,5′-цикло-АМФ до АМФ.
Некоторые вещества, обладающие ингибирующим действием на фосфодиэстеразу, (например, алкалоиды кофеин, теофиллин), способствуют сохранению и увеличению концентрации цикло-АМФ в клетке. Под действием этих веществ в организме продолжительность активации аденилатциклазной системы становится больше, т. е. усиливается действие гормона.
Кроме аденилатциклазной или гуанилатциклазной систем существует также механизм передачи информации внутри клетки-мишени с участием ионов кальция и инозитолтрифосфата.
Инозитолтрифосфат — это вещество, которое является производным сложного липида — инозитфосфатида. Оно образуется в результате действия специального фермента — фосфолипазы "С", который активируется в результате конформационных изменений внутриклеточного домена мембранного белка-рецептора.
Этот фермент гидролизует фосфоэфирную связь в молекуле фосфатидил-инозитол-4,5-бисфосфата и в результате образуются диацилглицерин и инозитолтрифосфат.
Известно, что образование диацилглицерина и инозитолтрифосфата приводит к увеличению концентрации ионизированного кальция внутри клетки. Это приводит к активации многих кальций-зависимых белков внутри клетки, в том числе активируются различные протеинкиназы. И здесь, как и при активации аденилатциклазной системы, одной из стадий передачи сигнала внутри клетки является фосфорилирование белков, которое в приводит к физиологическому ответу клетки на действие гормона.
В работе фосфоинозитидного механизма передачи сигналов в клетке-мишени принимает участие специальный кальций-связывающий белок — кальмодулин. Это низкомолекулярный белок (17 кДа), на 30 % состоящий из отрицательно заряженных аминокислот (Глу, Асп) и поэтому способный активно связывать Са+2. Одна молекула кальмодулина имеет 4 кальций-связывающих участка. После взаимодействия с Са+2 происходят конформационные изменения молекулы кальмодулина и комплекс "Са+2-кальмодулин" становится способным регулировать активность (аллостерически угнетать или активировать) многие ферменты — аденилатциклазу, фосфодиэстеразу, Са+2,Мg+2-АТФазу и различные протеинкиназы.
В разных клетках при воздействии комплекса "Са+2-кальмодулин" на изоферменты одного и того же фермента (например, на аденилатциклазу разного типа) в одних случаях наблюдается активация, а в других — ингибирование реакции образования цАМФ. Такие различные эффекты происходят потому, что аллостерические центры изоферментов могут включать в себя различные радикалы аминокислот и их реакция на действие комплекса Са+2-кальмодулин будет отличаться.
Таким образом, в роли "вторых посредников" для передачи сигналов от гормонов в клетках-мишенях могут быть:
комплекс "Са-кальмодулин";
диацилглицерин;
инозитолтрифосфат.
циклические нуклеотиды (ц-АМФ и ц-ГМФ);
Механизмы передачи информации от гормонов внутри клеток-мишеней с помощью перечисленных посредников имеют общие черты:
одним из этапов передачи сигнала является фосфорилирование белков;
прекращение активации происходит в результате специальных механизмов, инициируемых самими участниками процессов, — существуют механизмы отрицательной обратной связи.
Гормоны являются основными гуморальными регуляторами физиологических функций организма, и в настоящее время хорошо известны их свойства, процессы биосинтеза и механизмы действия.
Признаки, по которым гормоны отличаются от других сигнальных молекул следующие.
Синтез гормонов происходит в особых клетках эндокринной системы. При этом синтез гормонов является основной функцией эндокринных клеток.
Гормоны секретируются в кровь, чаще в венозную, иногда в лимфу. Другие сигнальные молекулы могут достигать клеток-мишеней без секреции в циркулирующие жидкости.
Телекринный эффект (или дистантное действие) — гормоны действуют на клетки-мишени на больщом расстоянии от места синтеза.
Гормоны являются высокоспецифичными веществами по отношению к клеткам-мишеням и обладают очень высокой биологической активностью.
Страница 4 из 9
Механизмы действия гормонов на клетки-мишени
В зависимости от строения гормона существуют два типа взаимодействия. Если молекула гормона липофильна, (например, стероидные гормоны), то она может проникать через липидный слой наружной мембраны клеток-мишеней. Если молекула имеет большие размеры или является полярной, то ее проникновение внутрь клетки невозможно. Поэтому для липофильных гормонов рецепторы находятся внутри клеток-мишеней, а для гидрофильных - рецепторы находятся в наружной мембране.
Для получения клеточного ответа на гормональный сигнал в случае гидрофильных молекул действует внутриклеточный механизм передачи сигнала. Это происходит с участием веществ, которых называют вторыми посредниками. Молекулы гормонов очень разнообразны по форме, а "вторые посредники" - нет.
Надежность передачи сигнала обеспечивает очень высокое сродство гормона к своему белку-рецептору.
Что такое посредники, которые участвуют во внутриклеточной передаче гуморальных сигналов?
Это циклические нуклеотиды (цАМФ и цГМФ), инозитолтрифосфат, кальций-связывающий белок - кальмодулин, ионы кальция, ферменты, участвующие в синтезе циклических нуклеотидов, а также протеинкиназы - ферменты фосфорилирования белков. Все эти вещества участвуют в регуляции активности отдельных ферментных систем в клетках-мишенях.
Разберем более подробно механизмы действия гормонов и внутриклеточных посредников.
Существует два главных способа передачи сигнала в клетки-мишени от сигнальных молекул с мембранным механизмом действия:
аденилатциклазная (или гуанилатциклазная) системы;
фосфоинозитидный механизм.
Прежде чем выяснить роль циклазной системы в механизме действия гормонов, рассмотрим определение этой системы. Система циклазная – это система, состоящая из содержащихся в клетке аденозинциклофосфата, аденилатциклазы и фосфодиэстеразы, регулирующая проницаемость клеточных мембран, участвует в регуляции многих обменных процессов живой клетки, опосредует действие некоторых гормонов. То есть роль циклазной системы заключается в том, что они являются вторыми посредниками в механизме действия гормонов.
Система «аденилатциклаза - цАМФ». Мембраны фермент аденилатциклаза может находиться в двух формах - активированной и неактивированной. Активация аденилатциклазы происходит под влиянием гормон-рецепторного комплекса, образование которого приводит к связыванию гуанилового нуклеотида (ГТФ) с особым регуляторным стимулирующим белком (GS-белок), после чего GS-белок вызывает присоединение магния к аденилатциклазе и ее активацию. Так действуют активизирующие аденилатциклазу гормоны глюкагон, тиреотропин, паратирин, вазопрессин, гонадотропин и др. Некоторые гормоны, напротив, подавляют аденилатциклазу (соматостатин, ангиотензин-П и др.).
Под влиянием аденилатциклазы из АТФ синтезируется цАМФ, вызывающий активацию протеинкиназ в цитоплазме клетки, обеспечивающих фосфорилирование многочисленных внутриклеточных белков. Это изменяет проницаемость мембран, т.е. вызывает типичные для гормона метаболические и, соответственно, функциональные сдвиги. Внутриклеточные эффекты цАМФ проявляются также во влиянии на процессы пролиферации, дифференцировки, на доступность мембранных рецепторных белков молекулам гормонов.
Система «гуанилатциклаза - цГМФ». Активация мембранной гуанилатциклазы происходит не под непосредственным влиянием гормон-рецепторного комплекса, а опосредованно через ионизированный кальций и оксидантные системы мембран. Так реализуют свои эффекты натрийуретический гормон предсердий - атриопептид, тканевой гормон сосудистой стенки. В большинстве тканей биохимические и физиологические эффекты цАМФ и цГМФ противоположны. Примерами могут служить стимуляция сокращений сердца под влиянием цАМФ и торможение их цГМФ, стимуляция сокращений гладких мышц кишечника цГМФ и подавление цАМФ.
Кроме аденилатциклазной или гуанилатциклазной систем существует также механизм передачи информации внутри клетки-мишени с участием ионов кальция и инозитолтрифосфата.
Инозитолтрифосфат - это вещество, которое является производным сложного липида - инозитфосфатида. Оно образуется в результате действия специального фермента - фосфолипазы "С", который активируется в результате конформационных изменений внутриклеточного домена мембранного белка-рецептора.
Этот фермент гидролизует фосфоэфирную связь в молекуле фосфатидил-инозитол-4,5-бисфосфата и в результате образуются диацилглицерин и инозитолтрифосфат.
Известно, что образование диацилглицерина и инозитолтрифосфата приводит к увеличению концентрации ионизированного кальция внутри клетки. Это приводит к активации многих кальций-зависимых белков внутри клетки, в том числе активируются различные протеинкиназы. И здесь, как и при активации аденилатциклазной системы, одной из стадий передачи сигнала внутри клетки является фосфорилирование белков, которое в приводит к физиологическому ответу клетки на действие гормона.
В работе фосфоинозитидного механизма передачи сигналов в клетке-мишени принимает участие специальный кальций-связывающий белок - кальмодулин. Это низкомолекулярный белок (17 кДа), на 30 % состоящий из отрицательно заряженных аминокислот (Глу, Асп) и поэтому способный активно связывать Са+2. Одна молекула кальмодулина имеет 4 кальций-связывающих участка. После взаимодействия с Са+2 происходят конформационные изменения молекулы кальмодулина и комплекс "Са+2-кальмодулин" становится способным регулировать активность (аллостерически угнетать или активировать) многие ферменты - аденилатциклазу, фосфодиэстеразу, Са+2,Мg+2-АТФазу и различные протеинкиназы.
В разных клетках при воздействии комплекса "Са+2-кальмодулин" на изоферменты одного и того же фермента (например, на аденилатциклазу разного типа) в одних случаях наблюдается активация, а в других - ингибирование реакции образования цАМФ. Такие различные эффекты происходят потому, что аллостерические центры изоферментов могут включать в себя различные радикалы аминокислот и их реакция на действие комплекса Са+2-кальмодулин будет отличаться.
Таким образом, в роли "вторых посредников" для передачи сигналов от гормонов в клетках-мишенях могут быть:
циклические нуклеотиды (ц-АМФ и ц-ГМФ);
комплекс "Са-кальмодулин";
диацилглицерин;
инозитолтрифосфат.
Механизмы передачи информации от гормонов внутри клеток-мишеней с помощью перечисленных посредников имеют общие черты:
одним из этапов передачи сигнала является фосфорилирование белков;
прекращение активации происходит в результате специальных механизмов, инициируемых самими участниками процессов, - существуют механизмы отрицательной обратной связи.
Гормоны являются основными гуморальными регуляторами физиологических функций организма, и в настоящее время хорошо известны их свойства, процессы биосинтеза и механизмы действия. Гормоны являются высокоспецифичными веществами по отношению к клеткам-мишеням и обладают очень высокой биологической активностью.
Стероидные гормоны являются относительно простыми органическими соединениями с небольшим молекулярным весом. О механизме их действия известно сейчас больше, чем о действии других гормонов. Скелет стероидных гормонов образован четырьмя углеводородными кольцами, и все разнообразие достигается за счет боковых групп – метальных, гидроксильных и др. Хотя сейчас известны десятки стероидных гормонов и их активных аналогов, общее число этих соединений, которые в принципе могут существовать, не превышает двухсот. Тем не менее в это число у позвоночных входят гормоны с совершенно различным действием – мужские половые гормоны (андростероны), женские половые гормоны (эстрогены), а также неполовые стероидные гормоны надпочечников – кортикостероиды.
Половые стероидные гормоны у позвоночных синтезируются в половых железах, и их синтез стимулируется гонадотропными гормонами гипофиза. У личинок насекомых стероидный гормон линьки – экдизон (экдистерон) синтезируется параторакальными железами.
Хорошей моделью действия женских половых стероидных гормонов (например, эстрадиола) служит синтез желточного белка ооцитов – вителлогенина в печени кур или овальбумина в яйцеводе цыплят. Для исследования начала синтеза вителлогенина часто используют петухов или самцов лягушек. У них в норме нет эстрогенов, вителлогенин не синтезируется и кодирующий его ген, очевидно, полностью не активен. Ho при введении эстрадиола синтез этого белка начинается и в печени самцов. Кроме генов вителлогенина, активируется также транскрипция рибосомных РНК и образование новых рибосом, в то время как активность других генов уменьшается. На новых мРНК и новых рибосомах интенсивно синтезируется вителлогенин и происходит его быстрый выход в кровяное русло. Однако, так как ооцитов у самцов нет, этот белок длительное время остается в плазме крови.
Введение эстрадиола молодым курочкам и даже цыплятам стимулирует дифференцировку клеток в их яйцеводах. Часть клеток эпителия яйцевода под влиянием эстрадиола дифференцируется в железистые клетки, в которых активируются гены овальбумина. Через несколько дней начинается синтез самого овальбумина.
В слюнных железах личинок дрозофилы или комара хирономуса (его личинки – это мотыль, живой корм для аквариумных рыб) действие стероидного гормона линьки – экдизона на активность генов можно видеть прямо под микроскопом. Политенные хромосомы значительно длиннее и толще обычных, и активные гены у них выглядят как утолщение хромосомы. Они называются пуфы. Уже через 5–10 мин после введения личинкам экдизона можно видеть появление нескольких новых пуфов (один – у хирономуса, два – у дрозофилы). Ho только через несколько часов у них возникает еще несколько десятков новых пуфов, появление которых характерно для личинки, вступившей в метаморфоз. Можно предполагать, что первые пуфы являются результатом прямого действия экдизона. Недавно при введении радиоактивного экдизона было показано, что он концентрируется в первых активирующихся пуфах. Более позднее включение остальных генов уже не требует прямого влияния экдизона и, вероятно, регулируется теми генами, которые активируются экдизоном в первые минуты. Механизм влияния «гена на ген» пора практически неизвестен, хотя такие влияния хорошо укладываются во многие схемы генной регуляции. Ингибиторы синтеза РНК подавляют включение второй очереди новых пуфов, но не препятствуют появлению первых пуфов.
Механизмы действия стероидных гормонов сейчас хорошо изучены. Эти гормоны проникают в клетки. В цитоплазме клеток‑мишеней находится специфический белок‑рецептор, который «опознает» тот гормон, к которому клетка компетентна, связывается с ним и образует гормон‑рецепторный комплекс. Такие комплексы мигрируют в ядро и связываются, как предполагают, с теми участками хромосом, где находятся гены, которые гормон в этих клетках активирует. Один и тот же стероидный гормон в разных видах клеток активирует разные гены, например эстрадиол активирует в печени гены вителлогенина, а в яйцеводе – гены овальбумина. Следовательно, эти клетки должны отличаться или своими рецепторами, или состоянием хромосом. Сейчас преобладает мнение, что рецепторы в разных видах клеток одинаковы. Если это так, то различными являются хромосомы. Предполагается, что в ядрах клеток‑мишеней на соответствующих генах находятся особые белки – акцепторы, с которыми может связываться гормон‑рецепторный комплекс и каким‑то (пока неясным) образом активировать данный ген.