К нерегулярным типам полового размножения можно отнести:
- партеногенетическое,
- гиногенетическое,
- андрогенетическое
размножение животных и растений.
Партеногенез - это развитие зародыша из неоплодотворенной яйцеклетки. Явление естественного партеногенеза свойственно низшим ракообразным, коловраткам, перепончатокрылым (пчелам, осам) и др. Известен он также у птиц (индейки). Партеногенез можно стимулировать искусственно, вызывая активацию неоплодотворенных яиц путем воздействия различными агентами. Различают партеногенез:
- соматический, или диплоидный,
- генеративный, или гаплоидный.
При соматическом партеногенезе яйцеклетка не претерпевает редукционного деления или если и претерпевает, то два гаплоидных ядра, сливаясь вместе, восстанавливают диплоидный набор хромосом (автокариогамия); таким образом в клетках тканей зародыша сохраняется диплоидный набор хромосом. При генеративном партеногенезе зародыш развивается из гаплоидной яйцеклетки. Например, у медоносной пчелы (Apis mellifera) трутни развиваются из неоплодотворенных гаплоидных яиц путем партеногенеза.
Гиногенез . Очень сходно с партеногенезом гиногенетическое размножение. В отличие от партеногенеза при гиногенезе участвуют сперматозоиды как стимуляторы развития яйцеклетки (псевдогамия), но оплодотворения (кариогамии) в этом случае не происходит; развитие зародыша осуществляется исключительно за счет женского ядра . Гиногенез обнаружен у круглых червей, живородящей рыбки Molliensia formosa, у серебряного карася (Platypoecilus) и у некоторых растений - лютика (Ranunculus auricomus), мятлика (род Роа pratensis) и др. Гиногенетическое развитие можно вызвать искусственно , если перед оплодотворением сперму или пыльцу облучить рентгеновыми лучами, обработать химическими веществами или подвергнуть действию высокой температуры. При этом разрушается ядро мужской гаметы и теряется способность к кариогамии, но сохраняется способность к активации яйца.
Явление гиногенетического размножения имеет большое значение для изучения наследственности , так как при этом потомство получает наследственную информацию только от матери . Таким образом, при бесполом размножении, партеногенезе и гиногенезе потомство должно быть сходно только с материнским организмом.
Андрогенез . Прямой противоположностью гиногенеза является андрогенез. При андрогенезе развитие яйца осуществляется только за счет мужских ядер и материнской цитоплазмы . Андрогенез может иметь место в тех случаях, когда материнское ядро почему-либо погибает до момента оплодотворения. Если в яйцеклетку попадает один сперматозоид, то развивающийся зародыш с гаплоидным набором хромосом оказывается нежизнеспособным или маложизнеспособным. Жизнеспособность андрогенных зигот нормализуется, если восстанавливается диплоидный набор хромосом.
Для нормального полового размножения характерны два процесса: образование мужских и женских гамет и формирование в результате их слияния зародыша, способного к развитию. Однако, в природе встречаются такие типы полового размножения, где один из этих процессов отсутствует. Это нерегулярные типы полового размножения: партеногенез, гиногенез, андрогенез .
Партеногенез : половое размножение, при котором зародыш развивается из неоплодотворенной яйцеклетки. Различают две формы партеногенеза: соматический (диплоидный) и генеративный (гаплоидный) .
Соматический партеногенез: яйцеклетка сохраняет диплоидный набор хромосом, поскольку в ней при мейозе не происходит редукционное деление, либо после него две гаплоидные клетки сливаются. Встречается у некоторых позвоночных (кавказская ящерица).
Генеративный партеногенез: мейоз протекает нормально, зародыш развивается из неоплодотворенной (гаплоидной яйцеклетки) Так размножаются некоторые виды членистоногих (у пчел развиваются самцы – трутни, у тлей – весеннее однополое поколение самок; у индеек – самцы).
У растений чаще всего встречается диплоидная форма партеногенеза апомиксис , разновидностью которого является апогамия (папоротники, цветковые). В последнем случае зародыш развивается из вегетативной диплоидной клетки спорофита. Апогамное размножение сочетается или чередуется с нормальным половым размножением (ястребинка, одуванчик, лапчатка и др.).
Гиногенез : вариант развития зародыша только из яйцеклетки. В отличие о партеногенеза, где участие мужской половой клетки полностью исключается, при гиногенезе сперматозоид проникает в яйцеклетку, однако слияния ядер не происходит, - сперматозоид лишь активирует яйцеклетку (круглые черви, некоторые рыбы, амфибии, некоторые высшие растения - лютик золотистый, мятлик луговой). Такое оплодотворение называется ложным или псевдогамией . При гиногенезе, как и партеногенезе, потомство получает наследственную информацию только от матери и тождественно ей по полу и признакам. Гиногенез можно вызвать искусственным путем, воздействуя на оплодотворенную яйцеклетку ионизирующими излучениями, химическими веществами, высокой температурой.
Андрогенез – развитие оплодотворенной яйцеклетки, у которой собственное ядро погибает еще до оплодотворения. Зародыш развивается за счет информации отцовского ядра и материнской цитоплазмы. Однако полноценным может быть лишь тогда, когда в яйцеклетку проникнут одновременно несколько сперматозоидов и если ядра двух гаплоидных спермиев сольются. Это создает условия для восстановления в клетке диплоидного набора хромосом. Потомство при таком размножении наследует признаки отцовского организма. Встречается редко у некоторых растений (табак, кукуруза) и животных (тутовый шелкопряд). Искусственно был вызван впервые в 40-х годах ХХ века Б. Л. Астауровым путем воздействия рентгеновским излучением на яйцеклетку тутового шелкопряда.
4. Половые клетки.
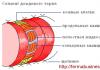
Яйцеклетка – женская генеративная (половая) клетка. Относительно крупная (от 60 мкм. до нескольких см.) неподвижная клетка обычно округлой формы; покрыта оболочкой, имеет большое количество цитоплазмы и ядро. Состав и структура цитоплазмы яйцеклеток являются видоспецифичными. В цитоплазме помимо типичных органоидов содержатся включения запасных питательных веществ в виде желтка. В ядрах клеток образуется много копий рибосомальных генов, и-РНК, обеспечивающих синтез жизненно важных белков будущего зародыша. Яйцеклетки разных организмов различаются количеством и характером распределения в них желтка. Различают несколько типов яйцеклеток. (рис…).
Изолецитальные (а) – относительно мелкие яйцеклетки с небольшим количеством равномерно распределенного желтка. Ядро располагается ближе к центру (черви, двустворчатые и брюхоногие моллюски, иглокожие, ланцетник).
Умеренно телолецитальные (б)– имеют диаметр около 1,5 – 2 мм, содержат среднее количество желтка, основная масса которого сосредоточена на вегетативном полюсе . На противоположном (анимальном ), где желтка мало, находится ядро яйцеклетки (земноводные, осетровые рыбы).
Резко телолецитальные – яйца крупные (10-15мм и более), содержат много желтка, занимающего почти весь объем цитоплазмы яйцеклетки. На анимальном полюсе находится зародышевый диск с активной, лишенной желтка цитоплазмой (некоторые рыбы, пресмыкающиеся, птицы, яйцекладущие млекопитающие).
Алецитальные – имеют микроскопически малые размеры (0,1-0,3 мм), практически лишены желтка (плацентарные млекопитающие, в том числе и человек).
1 – цитоплазма; 2 – ядро; 3 – блестящая оболочка; 4 – фолликулярные клетки.
Оболочки зрелой яйцеклетки делятся на первичные, вторичные и третичные.
Первичная оболочка (желточная) пронизана тяжами фолликулярных клеток, что при малом увеличении создает картину радиальной исчерченности, поэтому эта оболочка называется лучистым венцом (corona radiate); у млекопитающих она похожа на блестящий ободок и называется блестящая оболочка (zona pellucida).
Вторичная оболочка образуется за счет продуктов выделенияфолликулярных клеток, на стадии, когда яйцо находится в яичнике. Эта оболочка, хорион, имеется не у всех яиц. Ее особенность - микропиле - отверстие, через которое сперматозоид может проникнуть в яйцо.
Третичные оболочки формируются у ряда животных (амфибии, рептилии, птицы) за счет веществ, выделяемых железами яйцевода. У птиц они представлены белком, двумя слоями подскорлуповой оболочки и скорлупой.
 Сперматозоид
– мужская генеративная (половая) клетка. Обычно сперматозоиды очень мелкие (у человека – 50-70 мкм, у крокодила – 20 мкм); форма у разных видов варьирует, но большинство из них имеют головку, шейку и хвост. Головка содержит ядро с гаплоидным набором хромосом (1n1хр1с) и очень небольшое количество цитоплазмы. На переднем конце головки расположена акросома
– видоизмененный комплекс Гольджи, который содержит ферменты (гиалуронидазу и др.), растворяющие оболочки яйцеклетки при оплодотворении. В шейке находятся многочисленные митохондрии, которые образуют митохондриальную спираль, и центриоли. От шейки отрастает хвост, образованный микротрубочками и обеспечивающий подвижность сперматозоида. Разновидность сперматозоидов – клетки, лишенные хвоста, спермии
.
Сперматозоид
– мужская генеративная (половая) клетка. Обычно сперматозоиды очень мелкие (у человека – 50-70 мкм, у крокодила – 20 мкм); форма у разных видов варьирует, но большинство из них имеют головку, шейку и хвост. Головка содержит ядро с гаплоидным набором хромосом (1n1хр1с) и очень небольшое количество цитоплазмы. На переднем конце головки расположена акросома
– видоизмененный комплекс Гольджи, который содержит ферменты (гиалуронидазу и др.), растворяющие оболочки яйцеклетки при оплодотворении. В шейке находятся многочисленные митохондрии, которые образуют митохондриальную спираль, и центриоли. От шейки отрастает хвост, образованный микротрубочками и обеспечивающий подвижность сперматозоида. Разновидность сперматозоидов – клетки, лишенные хвоста, спермии
.
Гаметогенез.
Гаметогенез – процесс образования половых клеток, гамет, обычно протекает в половых железах (гонадах) . У высших организмов женские гаметы образуются в яичниках, а мужские – в семенниках. Образовавшиеся в результате овогенеза зрелые яйцеклетки, и в результате сперматогенеза зрелые сперматозоиды имеют гаплоидный набор хромосом (1n1хр1с).
В развитии половых клеток выделяют ряд стадий (или фаз).

Фаза размножения : характерна для ово- и сперматогенеза. Первичные половые клетки сперматогонии и овогонии размножаются в стенках семенника или яичка путем многократных митотических делений (2n1хр2с). У женщин размножение овогоний начинается в эмбриогенезе и завершается к 3-му году жизни. У мужчин фаза размножения начинается с наступлением половой зрелости и продолжается постоянно в течение всей жизни.
Фаза роста : овогонии и сперматогонии растут (увеличивается объем цитоплазмы, накопление веществ, необходимых для репликации ДНК, удвоения хромосом и дальнейшего деления); по завершении фазы роста они становятся овоцитами 1-го порядка и сперматоцитами 1-го порядка соответственно (2n2хр4с).
Фаза роста более выражена при овогенезе, поскольку овоциты 1 накапливают значительные количества питательных веществ. Рост овоцита 1-го порядка делят на два периода: малого и большого роста:
Малый рост – в этот период интенсивно выражены синтетические процессы, амплификация генов. Синтезированные и-РНК в основном используются развивающимся организмом после оплодотворения, и лишь небольшая доля в овогенезе;
Большой рост – изменений в ядре не происходит, объем плазмы увеличивается за счет отложения желтка (желток – вся совокупность питательных веществ клетки – белки, углеводы и жиры). Рост овоцита обеспечивается специальными механизмами питания с помощью фолликулярных клеток, соматических по своему происхождению, которые плотным кольцом охватывают овоцит. Фолликулярные клетки получают их кровеносных сосудов аминокислоты, белки, жиры и углеводы. Затем эти вещества попадают в овоцит. Запас веществ, формирующийся в период большого роста, расходуется после оплодотворения. Количество желтка зависит от длительности эмбрионального развития. Если вскоре после начала развития образуется личинка, способная питаться самостоятельно, желтка в яйце мало (у ланцетника маленькая личинка выходит через 4-5 дней после оплодотворения). Напротив, у птиц с крупным яйцом и большим количеством желтка развитие продолжается три недели и из яйцевых оболочек выходит в основном сформированный организм. Еще более продолжительный эмбриональный период у млекопитающих, но в этом случае зародыш питается за счет материнского организма и поэтому желтка в яйцеклетке очень мало. Увеличение объема яйцеклетки обусловлено увеличением объема цитоплазмы за счет накопления в ней большого количества нуклеотидов, РНК, белков. Резко возрастает объем ядра, т. к. в период роста в овоците образуется более 1 000 ядрышек, содержащих р-РНК.
Фаза созревания : созревание половых клеток происходит во время 1-го и 2-го мейотических делений. При сперматогенезе в результате мейоза 1 образуются два одинаковых спематоцита 2-го порядка (1n2хр2с), каждый из которых после мейоза 2 образует по две сперматиды (1n1хр1с). При овогенезе после первого мейотического деления образуется один овоцит 2-го порядка и одно направительное (редукционное ) тельце, которые после второго деления образуют соответственно овотиду и второе направительное тельце. Редукционные тельца содержат ядро и небольшое количество цитоплазмы; они «забирают» на себя излишки генетической информации и в дальнейшем погибают.
Деление созревания при овогенезе характеризуется рядом особенностей:
1. Профаза 1 мейоза проходит еще в эмбриональном периоде, а остальные события мейоза продолжаются после полового созревания.
2. Каждый месяц в одном из яичников половозрелой женщины созревает одна яйцеклетка; при этом завершается мейоз 1, образуется крупный овоцит 2-го порядка и маленькое полярное (направительное) тельце, которые вступают в меоз 2;
3. На стадии метафазы 2 овоцит 2-го порядка овулирует – выходит из яичника в брюшную полость, откуда попадает в яйцевод. Дальнейшее его созревание возможно лишь после слияния со сперматозоидом. Если оплодотворение не происходит овоцит 2 погибает и выводится из организма. В случае оплодотворения он завершает мейоз 2, образуя зрелую яйцеклетку – овотиду (1n1хр1с).
Таким образом, в результате фазы созревания из каждой диплоидной клетки, обладающей двухроматидными хромосомами (2n2хр2с), формируются гаплоидные клетки с однохроматидными хромосомами (1n1хр1с): при сперматогенезе – 4 сперматиды; при овогенезе – 1 овотида и 3 полярных тельца.
Фаза формирования : характерна только для сперматогенеза; в результате образуется подвижный сперматозоид с характерными чертами.
Таким образом гаметогенез завершается образованием генетически равноценных (1n1хр1с) половых клеток. Но эти яйцеклетка и сперматозоид неравноценны с точки зрения вклада в обеспечение развития будущего организма.
Функция сперматозоида – внесение генетической информации в яйцеклетку и активация ее развития. По своему строению сперматозоид специализирован для выполнения этой функции.
В яйцеклетке заложены все основные факторы, позволяющие организму развиваться , т. е. она специализирована для этой функции.
Сравнительная характеристика овогенеза и сперматогенеза
Оплодотворение.
Оплодотворение – процесс слияния сперматозоида и яйцеклетки, сопровождающийся объединением геномов отцовского и материнского организмов и завершающийся образованием зиготы. Сущность оплодотворения заключается в восстановлении двойного набора хромосом и в объединении наследственного материала обоих родителей, в результате чего потомство, соединяющее в себе полезные признаки отца и матери, более жизнеспособно: 1n1хр1с + 1n1хр1с = 2n1хр2с.
Встречу половых клеток обеспечивает процесс осеменения. Осеменение может быть наружным , когда половые продукты, содержащие сперматозоиды и яйцеклетки, выделяются в воду, где последние и встречаются (первично-водные животные – рыбы, земноводные), или внутренним , при котором самцы с помощью копулятивных органов вводят сперматозоиды в половые пути самки, где и происходит оплодотворение (членистоногие, пресмыкающиеся, птицы и млекопитающие).
Различают наружное оплодотворение, когда половые клетки сливаются вне организма, и внутреннее , когда половые клетки сливаются внутри половых путей самки. Кроме того, выделяют перекрестное оплодотворение, когда объединяются половые клетки разных особей, и самооплодотворение, которое происходит при слиянии гамет, продуцируемых одним и тем же организмом (гермафродиты у животных – плоские черви). В зависимости от числа сперматозоидов, оплодотворяющих одну яйцеклетку, выделяют моно- и полиспермию .
У млекопитающих и человека процесс оплодотворения происходит в маточной трубе, куда после овуляции попадают овоциты 2-го порядка и могут находиться многочисленные сперматозоиды.
Взаимодействие половых клеток делят на три фазы: дистантную, контактную и фазу взаимодействия после внедрения сперматозоида в яйцеклетку.
Дистантное взаимодействие обеспечивает встречу половых клеток после осеменения и ву некоторых организмов предохраняет яйцеклетку от проникновения в нее лишних сперматозоидов. Дистантное влияние яйцеклетки на сперматозоиды осуществляют гиногомоны -1 и гиногомоны- 2:
Гиногомоны-1 активирует действия сперматозоида, продлевают его подвижность;
Гиногомоны-2 (вещества белковой природы) вызывают склеивание сперматозоидов.
Влияние сперматозоидов носит несколько иной характер и обеспечивается андрогомонами-1 и аедрогомонами-2 :
Андрогомоны-1 (антогонисты гиногомона-1) выделяются во внешнюю среду сперматозоидами-лидерами и подавляют активность других сперматозоидов;
Андрогомоны-2 (белковые вещества, молекулы которого встроены в мембрану сперматозоида) обеспечивают склеивание сперматозоидов путем иммунной реакции с гиногомоном-2 (рис…)
Контактное взаимодействие
между сперматозоидом и яйцеклеткой осуществляется за счет акросомной реакции
. У млекопитающих она возникает под действием среды женских половых  органов и протекает без образования акросомного выроста. Фолликулярные клетки лучистого венца после овуляции сохраняются несколько часов. Поэтому после встречи сперматозоида с яйцеклеткой из акросомы освобождается фермент гиалуронидаза, который растворяет вещество, связывающее фолликулярные клетки вокруг яйца. Приблизившись к мембране яйца, сперматозоид сливается с его плазматической мембраной боковой поверхностью его головки. Это приводит к активации яйца с той стадии, с кото рой мейоз остановился. Реакция активации
заключается в переходе зрелого яйца из состояния покоя в состояние развития. В этот период повышается проницаемость мембраны для ионов К + и Са 2+ , активируется синтез липидов, белков, изменяется вязкость и другие коллоидные свойства белков яйца.
органов и протекает без образования акросомного выроста. Фолликулярные клетки лучистого венца после овуляции сохраняются несколько часов. Поэтому после встречи сперматозоида с яйцеклеткой из акросомы освобождается фермент гиалуронидаза, который растворяет вещество, связывающее фолликулярные клетки вокруг яйца. Приблизившись к мембране яйца, сперматозоид сливается с его плазматической мембраной боковой поверхностью его головки. Это приводит к активации яйца с той стадии, с кото рой мейоз остановился. Реакция активации
заключается в переходе зрелого яйца из состояния покоя в состояние развития. В этот период повышается проницаемость мембраны для ионов К + и Са 2+ , активируется синтез липидов, белков, изменяется вязкость и другие коллоидные свойства белков яйца.
Наиболее ярко активация яйцеклетки проявляется в кортикальной реакции : она начинается с места прикрепления сперматозоида к поверхности яйцеклетки (1). Под плазматической мембраной (2) располагаются кортикальные тельца (3), одетые своей собственной мембраной (содержат мукополисахариды, белки и другие вещества). Кортикальная реакция заключается в том, что после проникновения сперматозоида в яйцеклетку мембрана кортикальных телец слипается с плазматической мембраной. В месте слипания тельце раскрывается, его содержимое изливаеся и образует перивителлиновую жидкость , которая оттесняет желточную оболочку от поверхности ооплазмы. Желточная оболочка утолщается и становится ясно видимой и уже называется оболочкой оплодотворения.
Взаимодействие яйца и сперматозоида после его проникновения в яйцо заключается в основном в слиянии ядер (мужского и женского пронуклеусов ) с образованием диплоидного ядра - зиготы . На этом процесс оплодотворения заканчивается.
Вопросы для проверки самоподготовки :
1. Дать определение понятия «размножение», назвать основные его виды. В чем их отличия?
2. Перечислить способы бесполого размножения. В чем их суть? Привести примеры.
3. Назвать и охарактеризовать стадии гаметогенеза.
4. В чем заключается отличие сперматогенеза от овогенеза?
5. В чем заключаются отличия конъюгации и полового размножения?
6. Опишите строение сперматозоида и яйцеклетки.
7. Назовите и охарактеризуйте основные типы яйцеклеток..
8. Что такое осеменение? Назовите его виды. Привести примеры.
9. Что такое оплодотворение?
10. Опишите основные этапы оплодотворения.
Термины и понятия :
Акросома
Акросомная реакция
Андрогенез
Апогамия
Апомиксис
Бесполое размножение
Вегетативное размножение
Гетерогамия
Гиногенез
Изогамия
Клетки половые
Клетки соматические
Конъюгация
Копуляция
Кортикальная реакция
Микропиле
Оболочка оплодотворения
Овогоний
Оплодотворение
Осеменение
Партеногенез
Полиэбриония
Половое размножение
Псевдогамия
Размножение
Сингамия
Сперматида
Сперматогоний
Сперматозоид
Сперматоцит
Спорогония
Спорообразование
Фрагментация
Шизогония
У животных и растений встречаются так называемые нерегулярные типы полового размножения. Это прежде всего апомиксис (от греч. аро - из, от, mixis - смешение), т. е. половое размножение без оплодотворения. Апомиксис противоположен амфимиксису (от греч. amphi - с обеих сторон, mixis - смешение), т. е. половому размножению, происходящему путем слияния разнокачественных гамет. Синоним апомиксиса - партеногенез , т. е. девственное размножение (от греч. parihenos - девственница и genesis). Термин апомиксис чаще употребляют в отношении растений, а партеногенез - в отношении животных.
В 1958 г. И.С. Даревский описал популяции ящериц рода Lacerta , состоящие из одних самок и размножающиеся партеногенетичсски. Затем аналогичное явление было обнаружено у ящериц рода Cnemidaphorus. Оказалось, что у них перед мейозом в гониапьных клетках происходит эндомитотическое удвоение числа хромосом. Далее эти клетки проходят нормальный цикл мейоза, и в результате образуются диплоидные яйцеклетки, которые без оплодотворения дают начало новому поколению, состоящему только из самок.
С явлением апомиксиса у растений связаны драматические страницы в истории генетики. По неудачному совету К. Нэгели Г. Мендель после 1865 г. занялся проверкой открытых им закономерностей у ястребинок (Hieracium). Скрещивая разные виды этого растения, он обнаружил расщепление в F] и полное единообразие в F 2 . Получив этот результат, Г. Мендель опубликовал его в работе «О некоторых бастардах Hieracium , полученных искусственным оплодотворением» (1869) и бросил занятия гибридизацией. Только через 40 лет выяснилось, что Г. Мендель столкнулся с апомиктиче- ским размножением. Очевидно, выбранные формы были факультативно апомиктическими. В поколении, которое Г. Мендель считал первым гибридным, происходило расщепление исходно гетерозиготных форм. Теперь известно, что факультативные апомикты после гибридизации приобретают способность к устойчивому апомиксису.
Закономерности партеногенеза и хромосомного механизма определения пола у шелкопряда были использованы Б.Л. Астауровым для отбора наиболее продуктивных линий. Если извлечь из самок неоплодотворенные и не прошедшие мейоз яйца, то при прогревании их до 46 °С мейоз отсутствует. Эти яйца содержат Z- и Ж-хромосомы. Они развиваются партеноге- нетически и дают начало только самкам, т. к. у шелкопряда женский пол гетерогаметен. Таким образом можно быстро размножать ценный племенной материал.
Наряду с партеногенезом наблюдается и развитие яйцеклетки, активируемое сперматозоидом, не участвующим в оплодотворении. Мужской пронуклеус погибает, и организм развивается за счет женского пронуклеуса. Это явление называется гиногенезом и встречается у гермафродитных круглых червей и некоторых рыб.
Противоположность гиногенеза - андрогенез - развитие только за счет мужского пронуклеуса в случае гибели женского пронуклеуса. Гаплоидный андрогенез встречается очень редко. Развитие андрогенных особей до взрослого состояния наблюдали только у наездника Habrobracon и тутового шелкопряда.
У тутового шелкопряда при оплодотворении в яйцеклетку проникают несколько сперматозоидов, но ядро лишь одного из них сливается с ядром яйцеклетки, остальные погибают. Если неоплодотворенные яйцеклетки активировать температурным шоком, как описано выше, и облучить рентгеновскими лучами, то ядро яйцеклетки погибнет. Если далее такие энуклеированные яйца осеменить, то два мужских пронуклеуса, проникшие в яйцеклетку, сливаются между собой. За счет образовавшегося диплоидного ядра развивается зигота. Как показал Б.Л. Астауров, такие андрогенетические зиготы всегда превращаются в самцов, поскольку они несут две одинаковые половые хромосомы - ZZ. Получение чисто мужского потомства у шелкопряда экономически выгодно, т. к. самцы продуктивнее самок.
Половое размножение встречается в основном у высших организмов. Оно обеспечивает значительное генетическое разнообразие и, следовательно, большую фенотипическую изменчивость потомства; организмы получают большие эволюционные возможности, возникает материал для естественного отбора.
Помимо полового размножения, существует половой процесс. Суть его в том, что обмен генетической информацией между особями происходит, но без увеличения числа особей. Формированию гамет у многоклеточных предшествует мейоз. Половой процесс состоит в объединении наследственного материала от двух разных источников (родителей).
При половом размножении потомство генетически отличается от своих родителей, так как между родителями происходит обмен генетической информацией.
Основой полового размножения является мейоз. Родителями являются две особи – мужская и женская, они вырабатывают разные половые клетки. В этом проявляется половой диморфизм, который отражает различие задач, выполняемых при половом размножении мужским и женским организмами.
Половое размножение осуществляется через гаметы – половые клетки, имеющие гаплоидный набор хромосом и вырабатывающиеся в родительских организмах. Слияние родительских клеток приводит к образованию зиготы, из которой в дальнейшем образуется организм-потомок. Половые клетки образуются в гонадах – половых железах (в яичниках у самок и семенниках у самцов).
Процесс образования половых клеток называется гаметогенезом (овогенезом у самок и сперматогенезом у самцов).
Если мужские и женские гаметы образуются в организме одной особи, то ее называют гермафродитной. Гермафродитизм бывает истинный (особь имеет гонады обоих полов) и ложный гермафродитизм (особь имеет половые железы одного типа – мужского или женского, а наружные половые органы и вторичные половые признаки обоих полов).
Партеногенез (девственное размножение)
Виды партеногенеза:
1.облигатный (обязательный) партеногенез. Встречается в популяциях, состоящих исключительно из особей женского пола. При этом вероятность встречи разнополых особей минимальна
2.циклический (сезонный) партеногенез (у тлей, дафний, коловраток). Встречается в популяциях, которые исторически вымирали в больших количествах в определенное время года. У этих видов партеногенез сочетается с половым размножением. При этом в летнее время существуют только самки, которые откладывают два вида яиц – крупные и мелкие. Из крупных яиц партеногенетически появляются самки, а из мелких – самцы, которые оплодотворяют яйца, лежащие зимой на дне. Из них появляются исключительно самки;
3.факультативный (необязательный) партеногенез. Встречается у общественных насекомых (ос, пчел, муравьев). В популяции пчел из оплодотворенных яиц выходят самки (рабочие пчелы и царицы), из неоплодотворенных – самцы (трутни).
Гиногенез (у костистых рыб и некоторых земноводных). Сперматозоид проникает в яйцеклетку и лишь стимулирует ее развитие. Ядро сперматозоида при этом с ядром яйцеклетки не сливается и погибает, а источником наследственного материала для развития потомка служит ДНК ядра яйцеклетки.
Андрогенез. В развитии зародыша участвует мужское ядро, привнесенное в яйцеклетку, а ядро яйцеклетки при этом гибнет. Яйцеклетка дает лишь питательные вещества своей цитоплазмы.
Полиэмбриония . Зигота (эмбрион) делится на несколько частей бесполым способом, каждая из которых развивается в самостоятельный организм. Встречается у насекомых (наездников), броненосцев. У броненосцев клеточный материал первоначально одного зародыша на стадии бластулы равномерно разделяется между 4–8 зародышами, каждый из которых в дальнейшем дает полноценную особь.
У одноклеточных организмов выделяют две формы полового размножения – копуляцию и конъюгацию .
Наследование при апомиксисе . Законы доминирования и расщепления проявляются при половом размножении. Но в случае каких-либо отклонений от нормы в половом процессе происходят изменения характера расщепления. Рассмотрим нарушения менделевских закономерностей при нерегулярных типах полового размножения.
Уже Мендель приводит один из таких случаев. В опытах с ястребинкой (Hieracium) он столкнулся с большими трудностями в получении гибридов. У немногочисленных гибридов первого поколения он наблюдал не единообразие, а расщепление по окраске цветка. Второе же поколение оказалось единообразным, и отдельные растения целиком походили на гибриды F 1 , от которых они произошли. Таким образом, на этом объекте получились результаты, на первый взгляд противоположные законам доминирования и расщепления, открытым на горохе. Мендель не смог объяснить этот парадокс, так как он не знал особенностей размножения ястребинки (рис.7). Теперь известно, что у этого рода имеет место девственное размножение - апомиксис. Из яйцеклетки, которая образуется, минуя, мейоз , без оплодотворения возникает новый организм, генотипически полностью идентичный материнскому. Например, у растения генотипа Аа образуются такие же яйцеклетки Аа, в силу чего растения апомиктически размножающихся видов очень часто бывают гетерозиготными. Можно предположить, что в тех редких случаях, когда Менделю удалось осуществить гибридизацию, гетерозиготные растения дали расщепления. Когда гибридам F 1 была предоставлена возможность самоопыляться, они, размножаясь апомиктически, дали подобное себе единообразное потомство.

Рисунок 7. Окраска цветков ястребинки
Наследование при партеногенезе . В некоторых случаях при девственном размножении имеет место мейоз. Например, у пчел (Apis mellifera) из гаплоидных яйцеклеток при оплодотворении образуются самки, а без оплодотворения - самцы. В результате меняется и характер наследования признаков, как это можно видеть из следующей схемы:
Р ♀ аа х ♂А
серая желтый
 Гаметы а а А
Гаметы а а А
F ♂ а ♀ Аа
серый желтая
При скрещивании серой самки (аа) с желтым самцом (F) в F 1 получаются гибридные (Aа) желтые самки и партеногенетические (а) серые, подобные матерям самцы. Таким образом, при девственном размножении наследственная информация передается по материнской линии и партеногенетическое потомство (в данном случае самцы) похоже на мать.
Наследование при андрогенезе . В случае андрогенеза наблюдается противоположное явление, так как зигота развивается за счет цитоплазмы яйцеклетки и ядра сперматозоида и, следовательно, ее генотип будет определяться генотипом отца. Например, у самки шелкопряда (Bombyx mori), несущий доминантный признак коричневой окраски грены (АА), высокой температурой убивались ядра яйцеклеток. Такую самку скрещивали с самцом, имеющим рецессивный признак красной окраски грены (аа). В результате грена имела не коричневую окраску, как это было бы при нормальном оплодотворении у гибридных Аа зигот, а красную в соответствии с единственным геном а, полученным от отца. Таким образом, при андрогенезе потомство наследует отцовский признак.
В любых других случаях нарушений полового размножения будут наблюдаться отклонения от закономерностей наследования, открытых Менделем.
Необходимо подчеркнуть, что в подавляющем большинстве случаев при половом размножении на самых разных объектах и разнообразных признаках осуществляются открытые Менделем законы. Условия и причины, вызывающие нарушения проявления этих законов, связаны с аномалиями или изменениями характера полового процесса.
Наследование при бесполом размножении. При бесполом размножении, основой которого является равнонаследственное митотическое деление, характер наследования совсем иной. Рассмотрим такой пример. У земляники красная окраска ягоды (АА) неполно доминирует над белой (аа), у гетерозиготного растения (Аа) ягоды розовые. При самоопылении гетерозигота будет давать расщепление на красные, розовые и белые ягоды в отношении 1:2:1. Если же это гетерозиготное растение будет размножаться вегетативно, усами, то из отводок новые дочерние растения возникнут за счет митотических делений, т. е. каждая новая клетка и каждое дочернее растение будут иметь один и тот же генотип Аа, благодаря чему на всех растениях ягоды будут только розовые, как у исходной формы (рис.8).

Рисунок 8. Наследование окраски ягоды у земляники при самоопылении и вегетативном размножении: АА - красная; аа - белая; Ла - розовая окраска.
Таким образом, вегетативное размножение обеспечивает полное сходство потомков с родителями и единообразие особей в последовательных поколениях. Поэтому потомство одного вегетативно размножающегося растения всегда очень однородно, оно получило название клона.
Открытый Менделем метод анализа наследования отдельных пар признаков при моногибридном скрещивании позволил установить следующие закономерности:
1) признаки определяются константными наследственными задатками - генами,
2) при скрещивании в первом поколении наблюдается явление доминирования,
3) в потомстве гибрида (F 2) наблюдается расщепление в определенном количественном отношении.
Таким образом, своими исследованиями Мендель установил принципиально важное положение, а именно признаки (свойства) организма при скрещивании не исчезают в поколениях, а сохраняются. Это открытие явилось замечательным обоснованием учения Ч. Дарвина о происхождении видов путем естественного отбора. Оно позволило объяснить механизм, с помощью которого приспособительные свойства организмов не поглощаются скрещиванием, а сохраняются и могут накапливаться в поколениях под действием естественного отбора.