ЭМБРИОЛОГИЯ. Глава 21. ОСНОВЫ ЭМБРИОЛОГИИ ЧЕЛОВЕКА
ЭМБРИОЛОГИЯ. Глава 21. ОСНОВЫ ЭМБРИОЛОГИИ ЧЕЛОВЕКА
Эмбриология (от греч. embryon - зародыш, logos - учение) - наука о закономерностях развития зародышей.
Медицинская эмбриология изучает закономерности развития зародыша человека. Особое внимание обращается на эмбриональные источники и закономерные процессы развития тканей, метаболические и функциональные особенности системы мать-плацента-плод, критические периоды развития человека. Все это имеет большое значение для медицинской практики.
Знание эмбриологии человека необходимо всем врачам, особенно работающим в области акушерства и педиатрии. Это помогает в постановке диагноза при нарушениях в системе мать-плод, выявлении причин уродств и заболеваний детей после рождения.
В настоящее время знания по эмбриологии человека используются для раскрытия и ликвидации причин бесплодия, трансплантации фетальных органов, разработки и применения противозачаточных средств. В частности, актуальность приобрели проблемы культивирования яйцеклеток, экстракорпорального оплодотворения и имплантации зародышей в матку.
Процесс эмбрионального развития человека является результатом длительной эволюции и в определенной степени отражает черты развития других представителей животного мира. Поэтому некоторые ранние стадии развития человека очень сходны с аналогичными стадиями эмбриогенеза более низко организованных хордовых животных.
Эмбриогенез человека - часть его онтогенеза, включающая следующие основные стадии: I - оплодотворение и образование зиготы; II - дробление и образование бластулы (бластоцисты); III - гаструляцию - образование зародышевых листков и комплекса осевых органов; IV - гистогенез и органогенез зародышевых и внезародышевых органов; V - системогенез.
Эмбриогенез тесно связан с прогенезом и ранним постэмбриональным периодом. Так, развитие тканей начинается в эмбриональном периоде (эмбриональный гистогенез) и продолжается после рождения ребенка (постэмбриональный гистогенез).
21.1. ПРОГЕНЕЗ
Это период развития и созревания половых клеток - яйцеклеток и сперматозоидов. В результате проге-неза в зрелых половых клетках возникает гаплоидный набор хромосом, формируются структуры, обеспечивающие способность к оплодотворению и развитию нового организма. Подробно процесс развития половых клеток рассмотрен в главах, посвященных мужской и женской половым системам (см. главу 20).
Рис. 21.1. Строение мужской половой клетки:
I - головка; II - хвост. 1 - рецептор;
2 - акросома; 3 - «чехлик»; 4 - проксимальная центриоль; 5 - митохондрия; 6 - слой упругих фибрилл; 7 - аксоне-ма; 8 - терминальное кольцо; 9 - циркулярные фибриллы
Основные характеристики зрелых половых клеток человека
Мужские половые клетки
Сперматозоиды человека образуются в течение всего активного полового периода в больших количествах. Подробное описание сперматогенеза - см. главу 20.
Подвижность сперматозоидов обусловлена наличием жгутиков. Скорость движения сперматозоидов у человека равна 30-50 мкм/с. Целенаправленному движению способствуют хемотаксис (движение к химическому раздражителю или от него) и реотаксис (движение против тока жидкости). Через 30-60 мин после полового акта сперматозоиды обнаруживаются в полости матки, а через 1,5-2 ч - в дис-тальной (ампулярной) части маточной трубы, где происходят их встреча с яйцеклеткой и оплодотворение. Спермии сохраняют оплодотворяющую способность до 2 сут.
Строение. Мужские половые клетки человека - сперматозоиды, или спер-мии, длиной около 70 мкм, имеют головку и хвост (рис. 21.1). Плазмолемма сперматозоида в области головки содержит рецептор, с помощью которого происходит взаимодействие с яйцеклеткой.
Головка сперматозоида (caput spermatozoidi) включает небольшое плотное ядро с гаплоидным набором хромосом. Передняя половина ядра покрыта плоским мешочком, составляющим чехлик сперматозоида. В нем располагается акросома (от греч. асrоn - верхушка, soma - тело). Акросома содержит набор ферментов, среди которых важное место принадлежит гиалуронидазе и протеазам, способным растворять при оплодотворении оболочки, покрывающие яйцеклетку. Чехлик и акросома являются производными комплекса Гольджи.

Рис. 21.2. Клеточный состав эякулята человека в норме:
I - мужские половые клетки: А - зрелые (по Л. Ф. Курило и др.); Б - незрелые;
II - соматические клетки. 1, 2 - типичный сперматозоид (1 - анфас, 2 - профиль); 3-12 - наиболее часто встречающиеся формы атипии сперматозоидов; 3 - макроголовка; 4 - микроголовка; 5 - удлиненная головка; 6-7 - аномалия формы головки и акросомы; 8-9 - аномалия жгутика; 10 - двужгутиковый сперматозоид; 11 - сросшиеся головки (двухголовый сперматозоид); 12 - аномалия шейки сперматозоида; 13-18 - незрелые мужские половые клетки; 13-15 - первичные сперматоциты в профазе 1-го деления мейоза - пролептотена, пахитена, диплотена соответственно; 16 - первичный сперматоцит в метафазе мейоза; 17 - типичные сперматиды (а - ранняя; б - поздняя); 18 - атипичная двуядерная сперматида; 19 - эпителиальные клетки; 20-22 - лейкоциты
В ядре сперматозоида человека содержится 23 хромосомы, одна из которых является половой (X или Y), остальные - аутосомами. В 50 % сперматозоидов содержится Х-хромосома, в 50 % - Y-хромосома. Масса Х-хромосомы несколько больше массы Y-хромосомы, поэтому, видимо, сперматозоиды, содержащие Х-хромосому, менее подвижны, чем сперматозоиды, содержащие Y-хромосому.
За головкой имеется кольцевидное сужение, переходящее в хвостовой отдел.
Хвостовой отдел (flagellum) сперматозоида состоит из связующей, промежуточной, главной и терминальной частей. В связующей части (pars conjungens), или шейке (cervix), располагаются центриоли - проксимальная, прилежащая к ядру, и остатки дистальной центриоли, исчерченные колонны. Здесь начинается осевая нить (axonema), продолжающаяся в промежуточной, главной и терминальной частях.
Промежуточная часть (pars intermedia) содержит 2 центральных и 9 пар периферических микротрубочек, окруженных расположенными по спирали митохондриями (митохондриальное влагалище - vagina mitochondrialis). От микротрубочек отходят парные выступы, или «ручки», состоящие из другого белка - динеина, обладающего АТФ-азной активностью (см. главу 4). Динеин расщепляет АТФ, вырабатываемую митохондриями, и преобразует химическую энергию в механическую, за счет которой осуществляется движение спермия. В случае генетически обусловленного отсутствия динеина спермии оказываются обездвиженными (одна из форм стерильности мужчин).
Среди факторов, влияющих на скорость движения спермиев, большое значение имеют температура, рН среды и др.
Главная часть (pars principalis) хвоста по строению напоминает ресничку с характерным набором микротрубочек в аксонеме (9×2)+2, окруженных циркулярно ориентированными фибриллами, придающими упругость, и плазмолеммой.
Терминальная, или конечная, часть сперматозоида (pars terminalis) содержит аксонему, которая заканчивается разобщенными микротрубочками и постепенным уменьшением их числа.
Движения хвоста бичеобразные, что обусловлено последовательным сокращением микротрубочек от первой до девятой пары (первой считается пара микротрубочек, которая лежит в плоскости, параллельной двум центральным).
В клинической практике при исследовании спермы проводят подсчет различных форм сперматозоидов, подсчитывая их процентное содержание (спермиограмма).
По данным Всемирной организации здравоохранения (ВОЗ), нормальными характеристиками спермы человека являются следующие показатели: концентрация сперматозоидов - 20-200 млн/мл, содержание в эякуляте более 60 % нормальных форм. Наряду с последними в сперме человека всегда присутствуют аномальные - двужгутиковые, с дефектными размерами головки (макро- и микроформы), с аморфной головкой, со сросшимися
головками, незрелые формы (с остатками цитоплазмы в области шейки и хвоста), с дефектами жгутика.
В эякуляте здоровых мужчин преобладают типичные сперматозоиды (рис. 21.2). Количество различных видов атипичных сперматозоидов не должно превышать 30 %. Кроме того, встречаются незрелые формы половых клеток - сперматиды, сперматоциты (до 2 %), а также соматические клетки - эпителиоциты, лейкоциты.
Среди сперматозоидов в эякуляте живых клеток должно быть 75 % и более, а активно подвижных - 50 % и более. Установленные нормативные параметры необходимы для оценки отклонений от нормы при различных формах мужского бесплодия.
В кислой среде сперматозоиды быстро утрачивают способность к движению и оплодотворению.
Женские половые клетки
Яйцеклетки, или овоциты (от лат. ovum - яйцо), созревают в неизмеримо меньшем количестве, чем сперматозоиды. У женщины в течение полового цикла (24-28 дней) созревает, как правило, одна яйцеклетка. Таким образом, за детородный период образуются около 400 яйцеклеток.
Выход овоцита из яичника называется овуляцией (см. главу 20). Вышедший из яичника овоцит окружен венцом фолликулярных клеток, число которых достигает 3-4 тыс. Яйцеклетка имеет шаровидную форму, больший, чем у спермия, объем цитоплазмы, не обладает способностью самостоятельно передвигаться.
Классификация яйцеклеток основывается на признаках наличия, количества и распределения желтка (lecithos), представляющего собой белково-липидное включение в цитоплазме, используемое для питания зародыша. Различают безжелтковые (алецитальные), маложелтковые (олиголециталь-ные), среднежелтковые (мезолецитальные), многожелтковые (полилециталь-ные) яйцеклетки. Маложелтковые яйцеклетки подразделяются на первичные (у бесчерепных, например у ланцетника) и вторичные (у плацентарных млекопитающих и человека).
Как правило, в маложелтковых яйцеклетках желточные включения (гранулы, пластинки) распределены равномерно, поэтому они называются изолеци-тальными (греч. isos - равный). Яйцеклетка человека вторично изолецитального типа (как и у других млекопитающих животных) содержит небольшое количество желточных гранул, расположенных более или менее равномерно.
У человека наличие малого количества желтка в яйцеклетке обусловлено развитием зародыша в организме матери.
Строение. Яйцеклетка человека имеет диаметр около 130 мкм. К плазмо-лемме прилежат прозрачная (блестящая) зона (zona pellucida - Zp) и далее слой фолликулярных эпителиоцитов (рис. 21.3).
Ядро женской половой клетки имеет гаплоидный набор хромосом с X-половой хромосомой, хорошо выраженное ядрышко, в оболочке ядра много поровых комплексов. В период роста овоцита в ядре происходят интенсивные процессы синтеза иРНК, рРНК.

Рис. 21.3. Строение женской половой клетки:
1 - ядро; 2 - плазмолемма; 3 - фолликулярный эпителий; 4 - лучистый венец; 5 - кортикальные гранулы; 6 - желточные включения; 7 - прозрачная зона; 8 - рецептор Zp3
В цитоплазме развиты аппарат синтеза белка (эндоплазматическая сеть, рибосомы) и комплекс Гольджи. Количество митохондрий умеренно, они расположены около ядра, где идет интенсивный синтез желтка, клеточный центр отсутствует. Комплекс Гольджи на ранних стадиях развития располагается около ядра, а в процессе созревания яйцеклетки смещается на периферию цитоплазмы. Здесь располагаются производные этого комплекса - кортикальные гранулы (granula corticalia), число которых достигает 4000, а размеры 1 мкм. Они содержат гликозаминогликаны и различные ферменты (в том числе протеолитические), участвуют в кортикальной реакции, защищая яйцеклетку от полиспермии.
Из включений овоплазмы особого внимания заслуживают желточные гранулы, содержащие белки, фосфолипиды и углеводы. Каждая гранула желтка окружена мембраной, имеет плотную центральную часть, состоящую из фосфовитина (фосфопротеин), и более рыхлую периферическую часть, состоящую из липовителлина (липопротеин).
Прозрачная зона (zona pellucida - Zp) состоит из гликопротеинов и гли-козаминогликанов - хондроитинсерной, гиалуроновой и сиаловой кислот. Гликопротеины представлены тремя фракциями - Zpl, Zp2, Zp3. Фракции Zp2 и Zp3 образуют нити длиной 2-3 мкм и толщиной 7 нм, которые
соединены между собой с помощью фракции Zpl. Фракция Zp3 является рецептором спермиев, a Zp2 препятствует полиспермии. В прозрачной зоне содержатся десятки миллионов молекул гликопротеина Zp3, каждая из которых имеет более 400 аминокислотных остатков, соединенных со многими олигосахаридными ветвями. В образовании прозрачной зоны принимают участие фолликулярные эпителиоциты: отростки фолликулярных клеток проникают через прозрачную зону, направляясь к плазмолемме яйцеклетки. Плазмолемма яйцеклетки в свою очередь формирует микроворсинки, располагающиеся между отростками фолликулярных эпителиоцитов (см. рис. 21.3). Последние выполняют трофическую и защитную функции.
21.2. Эмбриогенез
Внутриутробное развитие человека продолжается в среднем 280 сут (10 лунных месяцев). Принято выделять три периода: начальный (1-я нед), зародышевый (2-8-я нед), плодный (с 9-й нед развития до рождения ребенка). К концу зародышевого периода завершается закладка основных эмбриональных зачатков тканей и органов.
Оплодотворение и образование зиготы
Оплодотворение (fertilisatio) - слияние мужской и женской половых клеток, в результате чего восстанавливается диплоидный набор хромосом, характерный для данного вида животных, и возникает качественно новая клетка - зигота (оплодотворенная яйцеклетка, или одноклеточный зародыш).
У человека объем эякулята - извергнутой спермы - в норме составляет около 3 мл. Для обеспечения оплодотворения общее количество сперматозоидов в сперме должно быть не менее 150 млн, а концентрация - 20- 200 млн/мл. В половых путях женщины после копуляции их число уменьшается по направлению от влагалища к ампулярной части маточной трубы.
В процессе оплодотворения различают три фазы: 1) дистантное взаимодействие и сближение гамет; 2) контактное взаимодействие и активизация яйцеклетки; 3) проникновение сперматозоида в яйцеклетку и последующее слияние - сингамия.
Первая фаза - дистантное взаимодействие - обеспечивается хемотаксисом - совокупностью специфических факторов, повышающих вероятность встречи половых клеток. Важную роль в этом играют гамоны - химические вещества, вырабатываемые половыми клетками (рис. 21.4). Например, яйцеклетки выделяют пептиды, способствующие привлечению сперматозоидов.
Сразу после эякуляции спермии не способны к проникновению в яйцеклетку до тех пор, пока не произойдет капацитация - приобретение спер-миями оплодотворяющей способности под действием секрета женских половых путей, которое длится 7 ч. В процессе капацитации с плазмолеммы спермия в области акросомы удаляются гликопротеины и протеины семенной плазмы, что способствует акросомальной реакции.

Рис. 21.4. Дистантное и контактное взаимодействие спермиев и яйцеклетки: 1 - сперматозоид и его рецепторы на головке; 2 - отделение углеводов с поверхности головки при капацитации; 3 - связывание рецепторов сперматозоида с рецепторами яйцеклетки; 4 - Zp3 (третья фракция гликопротеинов прозрачной зоны); 5 - плаз-молемма яйцеклетки; ГГI, ГГII - гиногамоны; АГI, АГII - андрогамоны; Гал - гли-козилтрансфераза; NАГ - N-ацетилглюкозамин
В механизме капацитации большое значение принадлежит гормональным факторам, прежде всего прогестерону (гормон желтого тела), активизирующему секрецию железистых клеток маточных труб. Во время капацитации происходят связывание холестерина плазмолеммы спермия альбуминами женских половых путей и обнажение рецепторов половых клеток. Оплодотворение происходит в ампулярной части маточной трубы. Оплодотворению предшествует осеменение - взаимодействие и сближение гамет (дистантное взаимодействие), обусловленное хемотаксисом.
Вторая фаза оплодотворения - контактное взаимодействие. Многочисленные спермии приближаются к яйцеклетке и вступают в контакт с ее оболочкой. Яйцеклетка начинает совершать вращательные движения вокруг своей оси со скоростью 4 оборота в минуту. Эти движения обусловлены биением хвостов сперматозоидов и продолжаются около 12 ч. Сперматозоиды при контакте с яйцеклеткой могут связывать десятки тысяч молекул гликопротеина Zp3. При этом отмечается запуск акросомальной реакции. Акросомальная реакция характеризуется повышением проницаемости плазмолеммы спермия к ионам Са 2 +, деполяризацией ее, что способствует слиянию плазмолеммы с передней мембраной акросомы. Прозрачная зона оказывается в непосредственном контакте с акросомальными ферментами. Ферменты разрушают ее, спермий проходит через прозрачную зону и

Рис. 21.5. Оплодотворение (по Вассерману с изменениями):
1-4 - стадии акросомной реакции; 5 - zona pellucida (прозрачная зона); 6 - периви-теллиновое пространство; 7 - плазматическая мембрана; 8 - кортикальная гранула; 8а - кортикальная реакция; 9 - проникновение спермия в яйцеклетку; 10 - зонная реакция
входит в перивителлиновое пространство, расположенное между прозрачной зоной и плазмолеммой яйцеклетки. Через несколько секунд изменяются свойства плазмолеммы яйцеклетки и начинается кортикальная реакция, а через несколько минут изменяются свойства прозрачной зоны (зонная реакция).
Инициация второй фазы оплодотворения происходит под влиянием суль-фатированных полисахаридов блестящей зоны, которые вызывают поступление ионов кальция и натрия в головку, спермия, замещение ими ионов калия и водорода и разрыв мембраны акросомы. Прикрепление спермия к яйцеклетке происходит под влиянием углеводной группы фракции гликопротеинов прозрачной зоны яйцеклетки. Рецепторы спермия представляют собой фермент гликозилтрансферазу, находящийся на поверхности акросомы головки, который

Рис. 21.6. Фазы оплодотворения и начало дробления (схема):
1 - овоплазма; 1а - кортикальные гранулы; 2 - ядро; 3 - прозрачная зона; 4 - фолликулярный эпителий; 5 - спермии; 6 - редукционные тельца; 7 - завершение митотического деления овоцита; 8 - бугорок оплодотворения; 9 - оболочка оплодотворения; 10 - женский пронуклеус; 11 - мужской пронуклеус; 12 - синкарион; 13 - первое митотическое деление зиготы; 14 - бластомеры
«узнает» рецептор женской половой клетки. Плазматические мембраны в месте контакта половых клеток сливаются, и происходит плазмогамия - объединение цитоплазм обеих гамет.
У млекопитающих при оплодотворении в яйцеклетку проникает лишь один сперматозоид. Такое явление называется моноспермией. Оплодотворению способствуют сотни других принимающих участие в осеменении сперматозоидов. Ферменты, выделяемые из акросом, - спермолизины (трипсин, гиалуронидаза) - разрушают лучистый венец, расщепляют гликозами-ногликаны прозрачной зоны яйцеклетки. Отделяющиеся фолликулярные эпителиоциты склеиваются в конгломерат, который вслед за яйцеклеткой перемещается по маточной трубе благодаря мерцанию ресничек эпителиальных клеток слизистой оболочки.

Рис. 21.7. Яйцеклетка и зигота человека (по Б. П. Хватову):
а - яйцеклетка человека после овуляции: 1 - цитоплазма; 2 - ядро; 3 - прозрачная зона; 4 - фолликулярные эпителиоциты, образующие лучистый венец; б - зигота человека в стадии сближения мужского и женского ядер (пронуклеусов): 1 - женское ядро; 2 - мужское ядро
Третья фаза. В овоплазму проникают головка и промежуточная часть хвостового отдела. После вхождения сперматозоида в яйцеклетку на периферии овоплазмы происходит уплотнение ее (зонная реакция) и образуется оболочка оплодотворения.
Кортикальная реакция - слияние плазмолеммы яйцеклетки с мембранами кортикальных гранул, в результате чего содержимое из гранул выходит в перивителлиновое пространство и воздействует на молекулы гликопро-теинов прозрачной зоны (рис. 21.5).
Вследствие этой зонной реакции молекулы Zp3 модифицируются и утрачивают способность быть рецепторами спермиев. Образуется оболочка оплодотворения толщиной 50 нм, препятствующая полиспермии - проникновению других спермиев.
Механизм кортикальной реакции включает приток ионов натрия через участок плазмолеммы сперматозоида, встроенный в плазмолемму яйцеклетки после завершения акросомальной реакции. В результате отрицательный мембранный потенциал клетки становится слабоположительным. Приток ионов натрия обусловливает высвобождение ионов кальция из внутриклеточных депо и увеличение его содержания в гиалоплазме яйцеклетки. Вслед за этим начинается экзоцитоз кортикальных гранул. Высвобождающиеся из них протеолитические ферменты разрывают связи между прозрачной зоной и плазмолеммой яйцеклетки, а также между спермиями и прозрачной зоной. Кроме того, выделяется гликопротеин, связывающий воду и привлекающий ее в пространство между плазмолеммой и прозрачной зоной. Вследствие этого формируется перивителлиновое пространство. Наконец,
выделяется фактор, способствующий затвердению прозрачной зоны и образованию из нее оболочки оплодотворения. Благодаря механизмам предотвращения полиспермии только одно гаплоидное ядро сперматозоида получает возможность слиться с одним гаплоидным ядром яйцеклетки, что приводит к восстановлению характерного для всех клеток диплоидного набора. Проникновение сперматозоида в яйцеклетку через несколько минут значительно усиливает процессы внутриклеточного обмена, что связано с активизацией ферментативных ее систем. Взаимодействие сперматозоидов с яйцеклеткой может быть заблокировано при помощи антител против веществ, входящих в прозрачную зону. На этом основании изыскиваются способы иммунологической контрацепции.
После сближения женского и мужского пронуклеусов, которое продолжается у млекопитающих около 12 ч, образуется зигота - одноклеточный зародыш (рис. 21.6, 21.7). На стадии зиготы выявляются презумптивные зоны (лат. presumptio - вероятность, предположение) как источники развития соответствующих участков бластулы, из которых в дальнейшем формируются зародышевые листки.
21.2.2. Дробление и образование бластулы
Дробление (fissio) - последовательное митотическое деление зиготы на клетки (бластомеры) без роста дочерних клеток до размеров материнской.
Образующиеся бластомеры остаются объединенными в единый организм зародыша. В зиготе образуется митотическое веретено между отдаляющи-

Рис. 21.8. Зародыш человека на ранних стадиях развития (по Гертигу и Рокку):
а - стадия двух бластомеров; б - бластоциста: 1 - эмбриобласт; 2 - трофобласт;
3 - полость бластоцисты

Рис. 21.9. Дробление, гаструляция и имплантация зародыша человека (схема): 1 - дробление; 2 - морула; 3 - бластоциста; 4 - полость бластоцисты; 5 - эмбрио-бласт; 6 - трофобласт; 7 - зародышевый узелок: а - эпибласт; б - гипобласт; 8 - оболочка оплодотворения; 9 - амниотический (эктодермальный) пузырек; 10 - внезародышевая мезенхима; 11 - эктодерма; 12 - энтодерма; 13 - цитотрофобласт; 14 - симпластотрофобласт; 15 - зародышевый диск; 16 - лакуны с материнской кровью; 17 - хорион; 18 - амниотическая ножка; 19 - желточный пузырек; 20 - слизистая оболочка матки; 21 - яйцевод
мися к полюсам центриолями, внесенными сперматозоидом. Пронуклеусы вступают в стадию профазы с формированием объединенного диплоидного набора хромосом яйцеклетки и сперматозоида.
Пройдя все остальные фазы митотического деления, зигота разделяется на две дочерние клетки - бластомеры (от греч. blastos - зачаток, meros - часть). Вследствие фактического отсутствия G 1 -периода, во время которого происходит рост клеток, образовавшихся в результате деления, клетки гораздо меньше материнской, поэтому и величина зародыша в целом в этот период независимо от числа составляющих его клеток не превышает величину исходной клетки - зиготы. Все это позволило назвать описываемый процесс дроблением (т. е. измельчением), а клетки, образующиеся в процессе дробления, - бластомерами.
Дробление зиготы человека начинается к концу первых суток и характеризуется как полное неравномерное асинхронное. В течение первых суток оно проис-
ходит медленно. Первое дробление (деление) зиготы завершается через 30 ч, в результате образуются два бластомера, покрытых оболочкой оплодотворения. За стадией двух бластомеров следует стадия трех бластомеров.
С первых же дроблений зиготы формируются два вида бластомеров - «темные» и «светлые». «Светлые», более мелкие, бластомеры дробятся быстрее и располагаются одним слоем вокруг крупных «темных», которые оказываются в середине зародыша. Из поверхностных «светлых» бластомеров в дальнейшем возникает трофобласт, связывающий зародыш с материнским организмом и обеспечивающий его питание. Внутренние, «темные», бластомеры формируют эмбриобласт, из которого образуются тело зародыша и внезародышевые органы (амнион, желточный мешок, аллантоис).
Начиная с 3 сут, дробление идет быстрее, и на 4-е сут зародыш состоит из 7-12 бластомеров. Через 50-60 ч образуется плотное скопление клеток - морула, а на 3-4-е сут начинается формирование бластоцисты - полого пузырька, заполненного жидкостью (см. рис. 21.8; рис. 21.9).
Бластоциста в течение 3 сут перемещается по маточной трубе к матке и через 4 сут попадает в полость матки. Бластоциста находится в полости матки в свободном виде (свободная бластоциста) в течение 2 сут (5-е и 6-е сут). К этому времени бластоциста увеличивается в размере благодаря росту числа бластомеров - клеток эмбриобласта и трофобла-ста - до 100 и вследствие усиленного всасывания трофобластом секрета маточных желез и активной выработки жидкости клетками трофобласта (см. рис. 21.9). Трофобласт первые 2 нед развития обеспечивает питание зародыша за счет продуктов распада материнских тканей (гистиотроф-ный тип питания),
Эмбриобласт располагается в виде узелка зародышевых клеток («зародышевый узелок»), который прикрепляется изнутри к трофобласту на одном из полюсов бластоцисты.
21.2.4. Имплантация
Имплантация (лат. implantatio - врастание, укоренение) - внедрение зародыша в слизистую оболочку матки.
Различают две стадии имплантации: адгезию (прилипание), когда зародыш прикрепляется к внутренней поверхности матки, и инвазию (погружение) - внедрение зародыша в ткани слизистой оболочки матки. На 7-е сут в трофобласте и эмбриобласте происходят изменения, связанные с подготовкой к имплантации. Бластоциста сохраняет оболочку оплодотворения. В трофобласте увеличивается количество лизосом с ферментами, обеспечивающими разрушение (лизис) тканей стенки матки и тем самым способствующими внедрению зародыша в толщу ее слизистой оболочки. Появляющиеся в трофобласте микроворсинки постепенно разрушают оболочку оплодотворения. Зародышевый узелок уплощается и превращается
в зародышевый щиток, в котором начинается подготовка к первой стадии гаструляции.
Имплантация продолжается около 40 ч (см. рис. 21.9; рис. 21.10). Одновременно с имплантацией начинается гаструляция (образование зародышевых листков). Это первый критический период развития.
В первой стадии трофобласт прикрепляется к эпителию слизистой оболочки матки, и в нем формируются два слоя - цитотрофобласт и симпластотро-фобласт. Во второй стадии симпластотрофобласт, продуцируя протеолити-ческие ферменты, разрушает слизистую оболочку матки. Формирующиеся при этом ворсинки трофобласта, внедряясь в матку, последовательно разрушают ее эпителий, затем подлежащую соединительную ткань и стенки сосудов, и трофобласт вступает в непосредственный контакт с кровью материнских сосудов. Образуется имплантационная ямка, в которой вокруг зародыша появляются участки кровоизлияний. Питание зародыша осуществляется непосредственно из материнской крови (гематотрофный тип питания). Из крови матери зародыш получает не только все питательные вещества, но и кислород, необходимый для дыхания. Одновременно в слизистой оболочке матки из клеток соединительной ткани, богатых гликогеном, происходит образование децидуальных клеток. После полного погружения зародыша в имплантационную ямку отверстие, образовавшееся в слизистой оболочке матки, заполняется кровью и продуктами разрушения тканей слизистой оболочки матки. В последующем дефект слизистой оболочки исчезает, эпителий восстанавливается путем клеточной регенерации.
Гематотрофный тип питания, сменяющий гистиотрофный, сопровождается переходом к качественно новому этапу эмбриогенеза - второй фазе гаструляции и закладке внезародышевых органов.
21.3. ГАСТРУЛЯЦИЯ И ОРГАНОГЕНЕЗ
Гаструляция (от лат. gaster - желудок) - сложный процесс химических и морфогенетических изменений, сопровождающийся размножением, ростом, направленным перемещением и дифференцировкой клеток, в результате чего образуются зародышевые листки: наружный (эктодерма), средний (мезодерма) и внутренний (энтодерма) - источники развития комплекса осевых органов и эмбриональных зачатков тканей.
Гаструляция у человека протекает в две стадии. Первая стадия (делами-нация) приходится на 7-е сут, а вторая стадия (иммиграция) - на 14-15-е сут внутриутробного развития.
При деламинации (от лат. lamina - пластинка), или расщеплении, из материала зародышевого узелка (эмбриобласта) образуются два листка: наружный листок - эпибласт и внутренний - гипобласт, обращенный в полость бла-стоцисты. Клетки эпибласта имеют вид псевдомногослойного призматического эпителия. Клетки гипобласта - мелкие кубические, с пенистой цито-

Рис. 21.10. Зародыши человека 7,5 и 11 сут развития в процессе имплантации в слизистую оболочку матки (по Гертигу и Рокку):
а - 7,5 сут развития; б - 11 сут развития. 1 - эктодерма зародыша; 2 - энтодерма зародыша; 3 - амниотический пузырек; 4 - внезародышевая мезенхима; 5 - цито-трофобласт; 6 - симпластотрофобласт; 7 - маточная железа; 8 - лакуны с материнской кровью; 9 - эпителий слизистой оболочки матки; 10 - собственная пластинка слизистой оболочки матки; 11 - первичные ворсинки
плазмой, формируют тонкий слой под эпибластом. Часть клеток эпибласта в дальнейшем образуют стенку амниотического пузырька, который начинает формироваться на 8-е сут. В области дна амниотического пузырька остается небольшая группа клеток эпибласта - материал, который пойдет на развитие тела зародыша и внезародышевых органов.
Вслед за деламинацией отмечается выселение клеток из наружного и внутреннего листков в полость бластоцисты, что знаменует формирование внезародышевой мезенхимы. К 11-м сут мезенхима подрастает к трофобласту и формируется хорион - ворсинчатая оболочка зародыша с первичными хориальными ворсинками (см. рис. 21.10).
Вторая стадия гаструляции происходит путем иммиграции (перемещения) клеток (рис. 21.11). Перемещение клеток происходит в области дна амниотического пузырька. Возникают клеточные потоки по направлению спереди назад, к центру и вглубь в результате размножения клеток (см. рис. 21.10). Это приводит к образованию первичной полоски. В головном конце первичная полоска утолщается, образуя первичный, или головной, узелок (рис. 21.12), откуда берет свое начало головной отросток. Головной отросток растет в краниальном направлении между эпи- и гипобластом и в дальнейшем дает начало развитию хорды зародыша, который определяет ось эмбриона, является основой развития костей осевого скелета. Вокруг хоры в будущем формируется позвоночный столб.
Клеточный материал, который перемещается из первичной полоски в пространство между эпибластом и гипобластом, располагается в виде мезо-дермальных крыльев парахордально. Часть клеток эпибласта внедряется в гипобласт, участвуя в образовании кишечной энтодермы. В результате зародыш приобретает трехслойное строение в виде плоского диска, состоящего из трех зародышевых листков: эктодермы, мезодермы и энтодермы.
Факторы, влияющие на механизмы гаструляции. Способы и скорость гастру-ляции определяются рядом факторов: дорсовентральным метаболическим градиентом, обусловливающим асинхронность размножения, дифференцировки и перемещения клеток; поверхностным натяжением клеток и межклеточными контактами, способствующими смещению групп клеток. Важную роль при этом играют индуктивные факторы. Согласно теории организационных центров, предложенной Г. Шпеманом, в определенных участках зародыша возникают индукторы (организующие факторы), которые оказывают индуцирующее влияние на другие участки зародыша, обусловливая их развитие в определенном направлении. Существуют индукторы (организаторы) нескольких порядков, действующих последовательно. Например, доказано, что организатор I порядка индуцирует развитие нервной пластинки из эктодермы. В нервной пластинке возникает организатор II порядка, способствующий превращению участка нервной пластинки в глазной бокал и т. п.
В настоящее время выяснена химическая природа многих индукторов (белки, нуклеотиды, стероиды и др.). Установлена роль щелевых контактов в межклеточных взаимодействиях. Под действием индукторов, исходящих из одной клетки, индуцируемая клетка, обладающая способностью специфического ответа, изменяет путь развития. Клетка, не подвергающаяся индукционному воздействию, сохраняет свои прежние потенции.
Дифференцировка зародышевых листков и мезенхимы начинается в конце 2-й - начале 3-й нед. Одна часть клеток преобразуется в зачатки тканей и органов зародыша, другая - во внезародышевые органы (см. главу 5, схему 5.3).

Рис. 21.11. Строение 2-недельного зародыша человека. Вторая стадия гаструляции (схема):
а - поперечный срез зародыша; б - зародышевый диск (вид со стороны амниоти-ческого пузырька). 1 - хориальный эпителий; 2 - мезенхима хориона; 3 - лакуны, заполненные материнской кровью; 4 - основание вторичной ворсины; 5 - амниоти-ческая ножка; 6 - амниотический пузырек; 7 - желточный пузырек; 8 - зародышевый щиток в процессе гаструляции; 9 - первичная полоска; 10 - зачаток кишечной энтодермы; 11 - желточный эпителий; 12 - эпителий амниотической оболочки; 13 - первичный узелок; 14 - прехордальный отросток; 15 - внезародышевая мезодерма; 16 - внезародышевая эктодерма; 17 - внезародышевая энтодерма; 18 - зародышевая эктодерма; 19 - зародышевая энтодерма

Рис. 21.12. Зародыш человека 17 сут («Крым»). Графическая реконструкция: а - эмбриональный диск (вид сверху) с проекцией осевых закладок и дефинитивной сердечно-сосудистой системой; б - сагиттальный (средний) срез через осевые закладки. 1 - проекция билатеральных закладок эндокарда; 2 - проекция билатеральных закладок перикардиального целома; 3 - проекция билатеральных закладок корпоральных кровеносных сосудов; 4 - амниотическая ножка; 5 - кровеносные сосуды в амниотической ножке; 6 - кровяные островки в стенке желточного пузырька; 7 - бухта аллантоиса; 8 - полость амниотического пузырька; 9 - полость желточного мешка; 10 - трофобласт; 11 - хордальный отросток; 12 - головной узелок. Условные обозначения: первичная полоска - штриховка вертикальная; первичный головной узелок обозначен крестами; эктодерма - без штриховки; энтодерма - линии; внезародышевая мезодерма - точки (по Н. П. Барсукову и Ю. Н. Шаповалову)
Дифференцировка зародышевых листков и мезенхимы, приводящая к появлению тканевых и органных зачатков, происходит неодновременно (гетерохронно), но взаимосвязанно (интегративно), в результате чего происходит формирование тканевых зачатков.
21.3.1. Дифференцировка эктодермы
При дифференцировке эктодермы образуются зародышевые части - кожная эктодерма, нейроэктодерма, плакоды, прехордальная пластинка, и вне-зародышевая эктодерма, являющаяся источником образования эпителиальной выстилки амниона. Меньшая часть эктодермы, расположенная над хордой (нейроэктодерма), дает начало дифференцировке нервной трубки и нервного гребня. Кожная эктодерма дает начало многослойному плоскому эпителию кожи (эпидермис) и ее производных, эпителию роговицы и конъюнктивы глаза, эпителию органов полости рта, эмали и кутикулы зубов, эпителию анального отдела прямой кишки, эпителиальной выстилке влагалища.
Нейруляция - процесс образования нервной трубки - протекает по времени неодинаково в различных частях зародыша. Замыкание нервной трубки начинается в шейном отделе, а затем распространяется кзади и несколько замедленнее в краниальном направлении, где формируются мозговые пузыри. Примерно на 25-е сут нервная трубка полностью замыкается, с внешней средой сообщаются только два незамкнувшихся отверстия на переднем и заднем концах - передний и задний невропоры (рис. 21.13). Задний невропор соответствует нейрокишечному каналу. Через 5-6 сут оба невропора зарастают. Из нервной трубки образуются нейроны и нейроглия головного и спинного мозга, сетчатки глаза и органа обоняния.
При смыкании боковых стенок нервных валиков и образовании нервной трубки появляется группа нейроэктодермальных клеток, образующихся в области соединения нейральной и остальной (кожной) эктодермы. Эти клетки, сначала располагающиеся в виде продольных рядов по обе стороны между нервной трубкой и эктодермой, образуют нервный гребень. Клетки нервного гребня способны к миграциям. В туловище одни клетки мигрируют в поверхностном слое дермы, другие - в вентральном направлении, образуя нейроны и нейроглию парасимпатических и симпатических узлов, хромаффинную ткань и мозговое вещество надпочечников. Часть клеток дифференцируется в нейроны и нейроглию спинномозговых узлов.
Из эпибласта выделяются клетки прехордальной пластинки, которая включается в состав головного отдела кишечной трубки. Из материала прехор-дальной пластинки развивается в дальнейшем многослойный эпителий переднего отдела пищеварительной трубки и ее производных. Кроме того, из прехордальной пластинки образуется эпителий трахеи, легких и бронхов, а также эпителиальная выстилка глотки и пищевода, производных жаберных карманов - тимуса и др.
По мнению А. Н. Бажанова, источником образования выстилки пищевода и дыхательных путей служит энтодерма головной кишки.

Рис. 21.13. Нейруляция у зародыша человека:
а - вид со спины; б - поперечные срезы. 1 - передний нейропор; 2 - задний ней-ропор; 3 - эктодерма; 4 - нервная пластинка; 5 - нервный желобок; 6 - мезодерма; 7 - хорда; 8 - энтодерма; 9 - нервная трубка; 10 - нервный гребень; 11 - головной мозг; 12 - спинной мозг; 13 - спинномозговой канал

Рис. 21.14. Зародыш человека на стадии образования туловищной складки и внезаро-дышевых органов (по П. Петкову):
1 - симпластотрофобласт; 2 - цитотрофобласт; 3 - внезародышевая мезенхима; 4 - место амниотической ножки; 5 - первичная кишка; 6 - полость амниона; 7 - эктодерма амниона; 8 - внезародышевая мезенхима амниона; 9 - полость желточного пузырька; 10 - энтодерма желточного пузырька; 11 - внезародышевая мезенхима желточного пузырька; 12 - аллантоис. Стрелками обозначено направление образования туловищной складки
В составе зародышевой эктодермы закладываются плакоды, являющиеся источником развития эпителиальных структур внутреннего уха. Из внезаро-дышевой эктодермы образуется эпителий амниона и пупочного канатика.
21.3.2. Дифференцировка энтодермы
Дифференцировка энтодермы приводит к образованию в теле зародыша энтодермы кишечной трубки и формированию внезародышевой энтодермы, формирующей выстилку желточного пузырька и аллантоиса (рис. 21.14).
Выделение кишечной трубки начинается с момента появления туловищной складки. Последняя, углубляясь, отделяет кишечную энтодерму будущей кишки от внезародышевой энтодермы желточного пузырька. В задней части зародыша в состав образующейся кишки входит и тот участок энтодермы, из которого возникает энтодермальный вырост аллантоиса.
Из энтодермы кишечной трубки развивается однослойный покровный эпителий желудка, кишечника и их желез. Кроме того, из энто-
дермы развиваются эпителиальные структуры печени и поджелудочной железы.
Внезародышевая энтодерма дает начало эпителию желточного мешка и аллантоиса.
21.3.3. Дифференцировка мезодермы
Этот процесс начинается на 3-й нед эмбриогенеза. Дорсальные участки мезодермы разделяются на плотные сегменты, лежащие по сторонам от хорды, - сомиты. Процесс сегментации дорсальной мезодермы и образования сомитов начинается в головной части зародыша и быстро распространяется в каудальном направлении.
У эмбриона на 22-е сут развития имеется 7 пар сегментов, на 25-е - 14, на 30-е - 30 и на 35-е - 43 -44 пары. В отличие от сомитов вентральные отделы мезодермы (спланхнотом) не сегментируются, а расщепляются на два листка - висцеральный и париетальный. Небольшой участок мезодермы, связывающий сомиты со спланхнотомом, разделяется на сегменты - сегментные ножки (нефрогонотом). На заднем конце зародыша сегментации этих отделов не происходит. Здесь взамен сегментных ножек располагается несегментированный нефрогенный зачаток (нефрогенный тяж). Из мезодермы зародыша развивается также парамезонефральный канал.
Сомиты дифференцируются на три части: миотом, дающий начало поперечнополосатой скелетной мышечной ткани, склеротом, являющийся источником развития костных и хрящевых тканей, а также дерматом, формирующий соединительнотканную основу кожи - дерму.
Из сегментных ножек (нефрогонотомов) развиваются эпителий почек, гонад и семявыводящих путей, а из парамезонефрального канала - эпителий матки, маточных труб (яйцеводов) и эпителий первичной выстилки влагалища.
Париетальный и висцеральный листки спланхнотома образуют эпителиальную выстилку серозных оболочек - мезотелий. Из части висцерального листка мезодермы (миоэпикардиальная пластинка) развиваются средняя и наружная оболочки сердца - миокард и эпикард, а также корковое вещество надпочечников.
Мезенхима в теле зародыша является источником формирования многих структур - клеток крови и кроветворных органов, соединительной ткани, сосудов, гладкой мышечной ткани, микроглии (см. главу 5). Из внезароды-шевой мезодермы развивается мезенхима, дающая начало соединительной ткани внезародышевых органов, - амниона, аллантоиса, хориона, желточного пузырька.
Соединительная ткань эмбриона и его провизорных органов характеризуется высокой гидрофильностью межклеточного вещества, богатством глико-заминогликанов в аморфном веществе. Соединительная ткань провизорных органов дифференцируется быстрее, чем в органных зачатках, что обусловлено потребностью в установлении связи зародыша с материнским организмом и
обеспечении их развития (например, плацента). Дифференцировка мезенхимы хориона наступает рано, но происходит не одновременно по всей поверхности. Наиболее активно процесс идет в области развития плаценты. Здесь же появляются и первые волокнистые структуры, которые играют важную роль в формировании и укреплении плаценты в матке. При развитии волокнистых структур стромы ворсин последовательно образуются сначала аргирофильные преколлагеновые волокна, а затем коллагеновые.
На 2-м мес развития в зародыше человека раньше всего начинается диф-ференцировка скелетогенной и кожной мезенхимы, а также мезенхимы стенки сердца и крупных кровеносных сосудов.
Артерии мышечного и эластического типа эмбрионов человека, а также артерии стволовых (якорных) ворсин плаценты и их разветвлений содержат десминотрицательные гладкие миоциты, обладающие свойством более быстрого сокращения.
На 7-й нед развития зародыша человека в кожной мезенхиме и мезенхиме внутренних органов появляются мелкие липидные включения, а позднее (8-9-я нед) происходит формирование жировых клеток. Вслед за развитием соединительной ткани сердечно-сосудистой системы дифференцируется соединительная ткань легких и пищеварительной трубки. Дифференцировка мезенхимы у зародышей человека (длиной 11-12 мм) на 2-м мес развития начинается с увеличения количества гликогена в клетках. В этих же участках возрастает активность фосфатаз, а в дальнейшем в ходе дифференцировки накапливаются гликопротеины, синтезируются РНК и белок.
Плодный период. Плодный период начинается с 9-й нед и характеризуется значительными морфогенетическими процессами, протекающими в организме как плода, так и матери (табл. 21.1).
Таблица 21.1. Краткий календарь внутриутробного развития человека (с дополнениями по Р. К. Данилову, Т. Г. Боровой, 2003)

Продолжение табл. 21.1

Продолжение табл. 21.1

Продолжение табл. 21.1

Продолжение табл. 21.1

Продолжение табл. 21.1

Продолжение табл. 21.1

Продолжение табл. 21.1

Окончание табл. 21.1

21.4. ВНЕЗАРОДЫШЕВЫЕ ОРГАНЫ
Внезародышевые органы, развивающиеся в процессе эмбриогенеза вне тела зародыша, выполняют многообразные функции, обеспечивающие рост и развитие самого зародыша. Некоторые из этих органов, окружающих зародыш, называют также зародышевыми оболочками. К этим органам относятся амнион, желточный мешок, аллантоис, хорион, плацента (рис. 21.15).
Источниками развития тканей внезародышевых органов являются троф-эктодерма и все три зародышевых листка (схема 21.1). Общие свойства тка-

Рис. 21.15. Развитие внезародышевых органов у зародыша человека (схема): 1 - амниотический пузырек; 1а - полость амниона; 2 - тело эмбриона; 3 - желточный мешок; 4 - внеэмбриональный целом; 5 - первичные ворсины хориона; 6 - вторичные ворсины хориона; 7 - стебелек аллантоиса; 8 - третичные ворсины хориона; 9 - аллан-тоис; 10 - пупочный канатик; 11 - гладкий хорион; 12 - котиледоны

Схема 21.1. Классификация тканей внезародышевых органов (по В. Д. Новикову, Г. В. Правоторову, Ю. И. Склянову)
ней внезародышевых органов и их отличия от дефинитивных сводятся к следующему: 1) развитие тканей является сокращенным и ускоренным; 2) соединительная ткань содержит мало клеточных форм, но много аморфного вещества, богатого гликозаминогликанами; 3) старение тканей внеза-родышевых органов происходит очень быстро - к концу внутриутробного развития.
21.4.1. Амнион
Амнион - временный орган, обеспечивающий водную среду для развития зародыша. Он возник в эволюции в связи с выходом позвоночных животных из воды на сушу. В эмбриогенезе человека он появляется на второй стадии гаструляции сначала как небольшой пузырек в составе эпибласта.
Стенка амниотического пузырька состоит из пласта клеток внезароды-шевой эктодермы и из внезародышевой мезенхимы, формирует его соединительную ткань.
Амнион быстро увеличивается, и к концу 7-й нед его соединительная ткань входит в контакт с соединительной тканью хориона. При этом эпителий амниона переходит на амниотическую ножку, превращающуюся позднее в пупочный канатик, и в области пупочного кольца смыкается с эпителиальным покровом кожи эмбриона.
Амниотическая оболочка образует стенку резервуара, заполненного амниотической жидкостью, в которой находится плод (рис. 21.16). Основная функция амниотической оболочки - выработка околоплодных вод, обеспечивающих среду для развивающегося организма и предохраняющих его от механического повреждения. Эпителий амниона, обращенный в его полость, не только выделяет околоплодные воды, но и принимает участие в обратном всасывании их. В амниотической жидкости поддерживаются до конца беременности необходимый состав и концентрация солей. Амнион выполняет также защитную функцию, предупреждая попадание в плод вредоносных агентов.
Эпителий амниона на ранних стадиях - однослойный плоский, образован крупными полигональными, тесно прилегающими друг к другу клетками, среди которых много митотически делящихся. На 3-м мес эмбриогенеза эпителий преобразуется в призматический. На поверхности эпителия имеются микроворсинки. В цитоплазме всегда содержатся небольшие капли липидов и гранулы гликогена. В апикальных частях клеток имеются различной величины вакуоли, содержимое которых выделяется в полость амниона. Эпителий амниона в области плацентарного диска однослойный призматический, местами многорядный, выполняет преимущественно секреторную функцию, в то время как эпителий вне-плацентарного амниона осуществляет в основном резорбцию околоплодных вод.
В соединительнотканной строме амниотической оболочки различают базальную мембрану, слой плотной волокнистой соединительной ткани и губчатый слой из рыхлой волокнистой соединительной ткани, связываю-

Рис. 21.16. Динамика взаимоотношений зародыша, внезародышевых органов и оболочек матки:
а - зародыш человека 9,5 нед развития (микрофотография): 1 - амнион; 2 - хорион; 3 - формирующаяся плацента; 4 - пуповина
щий амнион с хорионом. В слое плотной соединительной ткани можно выделить лежащую под базальной мембраной бесклеточную часть и клеточную часть. Последняя состоит из нескольких слоев фибробластов, между которыми находится густая сеть плотно прилежащих друг к другу тонких пучков коллагеновых и ретикулярных волокон, образующих решетку неправильной формы, ориентированную параллельно поверхности оболочки.
Губчатый слой образован рыхлой слизистой соединительной тканью с редкими пучками коллагеновых волокон, являющихся продолжением тех, которые залегают в слое плотной соединительной ткани, связывая амнион с хорионом. Связь эта очень непрочная, и поэтому обе оболочки легко отделить друг от друга. В основном веществе соединительной ткани много гликозаминогликанов.
21.4.2. Желточный мешок
Желточный мешок - наиболее древний в эволюции внезародышевый орган, возникший как орган, депонирующий питательные вещества (желток), необходимые для развития зародыша. У человека это рудиментарное образование (желточный пузырек). Он образован внезародышевой энтодермой и внезародышевой мезодермой (мезенхимой). Появившись на 2-й нед развития у человека, желточный пузырек в питании зародыша принимает

Рис. 21.16. Продолжение
б - схема: 1 - мышечная оболочка матки; 2 - decidua basalis; 3 - полость амниона; 4 - полость желточного мешка; 5 - внеэмбриональный целом (полость хориона); 6 - decidua capsularis; 7 - decidua parietalis; 8 - полость матки; 9 - шейки матки; 10 - эмбрион; 11 - третичные ворсинки хориона; 12 - аллантоис; 13 - мезенхима пупочного канатика: а - кровеносные сосуды ворсины хориона; б - лакуны с материнской кровью (по Гамильтону, Бойду и Моссману)
участие очень недолго, так как с 3-й нед развития устанавливается связь плода с материнским организмом, т. е. гематотрофное питание. Желточный мешок позвоночных является первым органом, в стенке которого развиваются кровяные островки, формирующие первые клетки крови и первые кровеносные сосуды, обеспечивающие у плода перенос кислорода и питательных веществ.
По мере образования туловищной складки, приподнимающей зародыш над желточным пузырьком, формируется кишечная трубка, при этом желточный пузырек отделяется от тела зародыша. Связь зародыша с желточным пузырьком остается в виде полого канатика, называемого желточным стебельком. В качестве кроветворного органа желточный мешок функционирует до 7-8-й нед, а затем подвергается обратному развитию и остается в составе пупочного канатика в виде узкой трубочки, служащей проводником кровеносных сосудов к плаценте.
21.4.3. Аллантоис
Аллантоис представляет собой небольшой пальцевидный отросток в кау-дальном отделе зародыша, врастающий в амниотическую ножку. Он является производным желточного мешка и состоит из внезародышевой энтодермы и висцерального листка мезодермы. У человека аллантоис не достигает значительного развития, но его роль в обеспечении питания и дыхания зародыша все же велика, так как по нему к хориону растут сосуды, располагающиеся в пупочном канатике. Проксимальная часть аллантоиса располагается вдоль желточного стебелька, а дистальная, разрастаясь, врастает в щель между амнионом и хорионом. Это орган газообмена и выделения. По сосудам аллантоиса доставляется кислород, а в аллантоис выделяются продукты обмена веществ зародыша. На 2-м мес эмбриогенеза аллантоис редуцируется и превращается в тяж клеток, который вместе с редуцированным желточным пузырьком входит в состав пупочного канатика.
21.4.4. Пупочный канатик
Пупочный канатик, или пуповина, представляет собой упругий тяж, соединяющий зародыш (плод) с плацентой. Он покрыт амниотической оболочкой, окружающей слизистую соединительную ткань с кровеносными сосудами (две пупочные артерии и одна вена) и рудиментами желточного пузырька и аллантоиса.
Слизистая соединительная ткань, получившая название «вартонова студня», обеспечивает упругость канатика, предохраняет пупочные сосуды от сжатия, обеспечивая тем самым непрерывное снабжение эмбриона питательными веществами, кислородом. Наряду с этим она препятствует проникновению вредоносных агентов из плаценты к эмбриону внесосудистым путем и таким образом выполняет защитную функцию.
Иммуноцитохимическими методами установлено, что в кровеносных сосудах пупочного канатика, плаценты и эмбриона существуют гетерогенные гладкие мышечные клетки (ГМК). В венах в отличие от артерий обнаружены десминположительные ГМК. Последние обеспечивают медленные тонические сокращения вен.
21.4.5. Хорион
Хорион, или ворсинчатая оболочка, появляется впервые у млекопитающих, развивается из трофобласта и внезародышевой мезодермы. Первоначально трофобласт представлен слоем клеток, образующих первичные ворсинки. Они выделяют протеолитические ферменты, с помощью которых разрушается слизистая оболочка матки и осуществляется имплантация. На 2-й нед трофобласт приобретает двухслойное строение в связи с формированием в нем внутреннего клеточного слоя (цитотрофобласт) и симпластического наружного слоя (симпластотрофобласт), который является производным клеточного слоя. Появляющаяся по периферии эмбриобласта внезародыше-вая мезенхима (у человека на 2-3-й нед развития) подрастает к трофобла-сту и образует вместе с ним вторичные эпителиомезенхимальные ворсинки. С этого времени трофобласт превращается в хорион, или ворсинчатую оболочку (см. рис. 21.16).
В начале 3-й нед в ворсинки хориона врастают кровеносные капилляры и формируются третичные ворсинки. Это совпадает с началом гема-тотрофного питания зародыша. Дальнейшее развитие хориона связано с двумя процессами - разрушением слизистой оболочки матки вследствие протеолитической активности наружного (симпластического) слоя и развитием плаценты.
21.4.6. Плацента
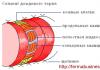
Плацента (детское место) человека относится к типу дискоидальных гемохориальных ворсинчатых плацент (см. рис. 21.16; рис. 21.17). Это важный временный орган с многообразными функциями, которые обеспечивают связь плода с материнским организмом. Вместе с тем плацента создает барьер между кровью матери и плода.
Плацента состоит из двух частей: зародышевой, или плодной (pars fetalis), и материнской (pars materna). Плодная часть представлена ветвистым хорионом и приросшей к хориону изнутри амниотической оболочкой, а материнская - видоизмененной слизистой оболочкой матки, отторгающейся при родах (decidua basalis).
Развитие плаценты начинается на 3-й нед, когда во вторичные ворсины начинают врастать сосуды и образовываться третичные ворсины, и заканчивается к концу 3-го мес беременности. На 6-8-й нед вокруг сосудов

Рис. 21.17. Плацента гемохориального типа. Динамика развития ворсин хориона: а - строение плаценты (стрелками указана циркуляция крови в сосудах и в одной из лакун, где удалена ворсинка): 1 - эпителий амниона; 2 - хориальная пластинка; 3 - ворсинка; 4 - фибриноид; 5 - желточный пузырек; 6 - пупочный канатик; 7 - перегородка плаценты; 8 - лакуна; 9 - спиральная артерия; 10 - базальный слой эндометрия; 11 - миометрий; б - строение первичной ворсины трофобласта (1-я нед); в - строение вторичной эпителиально-мезенхимальной ворсины хориона (2-я нед); г - строение третичной ворсины хориона - эпителиально-мезенхимальной с кровеносными сосудами (3-я нед); д - строение ворсины хориона (3-й мес); е - строение ворсин хориона (9-й мес): 1 - межворсинчатое пространство; 2 - микроворсинки; 3 - симпластотрофобласт; 4 - ядра симпластотрофобласта; 5 - цито-трофобласт; 6 - ядро цитотрофобласта; 7 - базальная мембрана; 8 - межклеточное пространство; 9 - фибробласт; 10 - макрофаги (клетки Кащенко-Гофбауэра); 11 - эндотелиоцит; 12 - просвет кровеносного сосуда; 13 - эритроцит; 14 - базальная мембрана капилляра (по Э. М. Швирсту)
дифференцируются элементы соединительной ткани. В дифференцировке фибробластов и синтезе ими коллагена важную роль играют витамины А и С, без достаточного поступления которых в организм беременной нарушается прочность связи зародыша с материнским организмом и создается угроза самопроизвольного аборта.
В основном веществе соединительной ткани хориона содержится значительное количество гиалуроновой и хондроитинсерной кислот, с которыми связана регуляция проницаемости плаценты.
При развитии плаценты происходят разрушение слизистой оболочки матки, обусловленное протеолитической активностью хориона, и смена гистиотрофного питания на гематотрофное. Это означает, что ворсины хориона омываются кровью матери, излившейся из разрушенных сосудов эндометрия в лакуны. Однако кровь матери и плода в нормальных условиях никогда не смешивается.
Гематохориальный барьер, разделяющий оба кровотока, состоит из эндотелия сосудов плода, окружающей сосуды соединительной ткани, эпителия хориальных ворсин (цитотрофобласт и симпластотрофобласт), а кроме того, из фибриноида, который местами покрывает ворсины снаружи.
Зародышевая, или плодная, часть плаценты к концу 3-го мес представлена ветвящейся хориальной пластинкой, состоящей из волокнистой (коллаге-новой) соединительной ткани, покрытой цито- и симпластотрофобластом (многоядерная структура, покрывающая редуцирующийся цитотрофо-бласт). Ветвящиеся ворсины хориона (стволовые, якорные) хорошо развиты лишь со стороны, обращенной к миометрию. Здесь они проходят через всю толщу плаценты и своими вершинами погружаются в базальную часть разрушенного эндометрия.
Хориальный эпителий, или цитотрофобласт, на ранних стадиях развития представлен однослойным эпителием с овальными ядрами. Эти клетки размножаются митотическим путем. Из них развивается симпластотрофобласт.
В симпластотрофобласте содержится большое количество различных про-теолитических и окислительных ферментов (АТФ-азы, щелочная и кислая

Рис. 21.18. Срез ворсины хориона 17-суточного зародыша человека («Крым»). Микрофотография:
1 - симпластотрофобласт; 2 - цитотрофобласт; 3 - мезенхима хориона (по Н. П. Барсукову)
фосфатазы, 5-нуклеотидазы, ДПН-диафоразы, глюкозо-6-фосфатдегид-рогеназы, альфа-ГФДГ, сукцинатдегидрогеназа - СДГ, цитохромоксидаза - ЦО, моноаминоксидаза - МАО, неспецифические эстеразы, ЛДГ, НАД- и НАДФ-диафоразы и др. - всего около 60), что связано с его ролью в обменных процессах между организмом матери и плода. В цитотрофобласте и в симпласте выявляются пиноцитозные пузырьки, лизосомы и другие орга-неллы. Начиная со 2-го мес, хориальный эпителий истончается и постепенно заменяется симпластотрофобластом. В этот период симпластотрофобласт по толщине превосходит цитотрофобласт. На 9-10-й нед симпласт истончается, а количество ядер в нем увеличивается. На поверхности симпласта, обращенной в лакуны, появляются многочисленные микроворсинки в виде щеточной каемки (см. рис. 21.17; рис. 21.18, 21.19).
Между симпластотрофобластом и клеточным трофобластом имеются ще-левидные субмикроскопические пространства, доходящие местами до ба-зальной мембраны трофобласта, что создает условия для двустороннего проникновения трофических веществ, гормонов и др.
Во второй половине беременности и, особенно, в конце нее трофобласт сильно истончается и ворсины покрываются фибриноподобной оксифиль-ной массой, являющейся продуктом свертывания плазмы и распада трофо-бласта («фибриноид Лангханса»).
С увеличением срока беременности уменьшается количество макрофагов и коллагенпродуцирующих дифференцированных фибробластов, появля-

Рис. 21.19. Плацентарный барьер на 28-й нед беременности. Электронная микрофотография, увеличение 45 000 (по У. Ю. Яцожинской):
1 - симпластотрофобласт; 2 - цитотрофобласт; 3 - базальная мембрана трофобласта; 4 - базальная мембрана эндотелия; 5 - эндотелиоцит; 6 - эритроцит в капилляре
ются фиброциты. Количество коллагеновых волокон, хотя и нарастает, но до конца беременности в большинстве ворсин остается незначительным. Большая часть стромальных клеток (миофибробластов) характеризуется увеличенным содержанием цитоскелетных сократительных белков (вимен-тин, десмин, актин и миозин).
Структурно-функциональной единицей сформированной плаценты является котиледон, образованный стволовой («якорной») ворсиной и ее
вторичными и третичными (конечными) разветвлениями. Общее количество котиледонов в плаценте достигает 200.
Материнская часть плаценты представлена базальной пластинкой и соединительнотканными септами, отделяющими котиледоны друг от друга, а также лакунами, заполненными материнской кровью. В местах контакта стволовых ворсин с отпадающей оболочкой встречаются также трофобла-стические клетки (периферический трофобласт).
На ранних стадиях беременности ворсины хориона разрушают ближайшие к плоду слои основной отпадающей оболочки матки, и на их месте образуются заполненные материнской кровью лакуны, в которые свободно свисают ворсины хориона.
Глубокие неразрушенные части отпадающей оболочки вместе с трофо-бластом образуют базальную пластинку.
Базальный слой эндометрия (lamina basalis) - соединительная ткань слизистой оболочки матки, содержащая децидуальные клетки. Эти крупные, богатые гликогеном клетки соединительной ткани расположены в глубоких слоях слизистой оболочки матки. Они имеют четкие границы, округлые ядра и оксифильную цитоплазму. В течение 2-го мес беременности децидуальные клетки значительно укрупняются. В их цитоплазме, кроме гликогена, выявляются липиды, глюкоза, витамин С, железо, неспецифические эстеразы, дегидрогеназа янтарной и молочной кислот. В базальной пластинке, чаще в месте прикрепления ворсин к материнской части плаценты, встречаются скопления клеток периферического цитотрофобласта. Они напоминают децидуальные клетки, но отличаются более интенсивной базо-филией цитоплазмы. Аморфная субстанция (фибриноид Рора) находится на поверхности базальной пластинки, обращенной к хориальным ворсинам. Фибриноид играет существенную роль в обеспечении иммунологического гомеостаза в системе мать-плод.
Часть основной отпадающей оболочки, расположенной на границе ветвистого и гладкого хориона, т. е. по краю плацентарного диска, при развитии плаценты не разрушается. Плотно прирастая к хориону, она образует замыкающую пластинку, препятствующую истечению крови из лакун плаценты.
Кровь в лакунах непрерывно циркулирует. Она поступает из маточных артерий, входящих сюда из мышечной оболочки матки. Эти артерии идут по плацентарным перегородкам и открываются в лакуны. Материнская кровь оттекает от плаценты по венам, берущим начало от лакун крупными отверстиями.
Формирование плаценты заканчивается в конце 3-го мес беременности. Плацента обеспечивает питание, тканевое дыхание, рост, регуляцию образовавшихся к этому времени зачатков органов плода, а также его защиту.
Функции плаценты. Основные функции плаценты: 1) дыхательная; 2) транспорт питательных веществ; воды; электролитов и иммуноглобулинов; 3) выделительная; 4) эндокринная; 5) участие в регуляции сокращения миометрия.
Дыхание плода обеспечивается за счет кислорода, присоединенного к гемоглобину материнской крови, который путем диффузии поступает через плаценту в кровь плода, где он соединяется с фетальным гемоглобином
(HbF). Связанный с фетальным гемоглобином СО 2 в крови плода также диффундирует через плаценту, поступает в кровь матери, где соединяется с материнским гемоглобином.
Транспорт всех питательных веществ, необходимых для развития плода (глюкоза, аминокислоты, жирные кислоты, нуклеотиды, витамины, минеральные вещества), происходит из крови матери через плаценту в кровь плода, и, наоборот, из крови плода в кровь матери поступают продукты обмена веществ, выводимые из его организма (выделительная функция). Электролиты и вода проходят через плаценту путем диффузии и с помощью пиноцитоза.
В транспорте иммуноглобулинов участвуют пиноцитозные везикулы симпластотрофобласта. Поступивший в кровь плода иммуноглобулин пассивно иммунизирует его от возможного действия бактериальных антигенов, которые могут поступать при заболеваниях матери. После рождения материнский иммуноглобулин разрушается и заменяется вновь синтезируемым в организме ребенка при действии на него бактериальных антигенов. Через плаценту в околоплодные воды проникают IgG, IgA.
Эндокринная функция является одной из наиболее важных, так как плацента обладает способностью синтезировать и секретировать ряд гормонов, обеспечивающих взаимодействие зародыша и материнского организма на протяжении всей беременности. Местом продукции плацентарных гормонов являются цитотрофобласт и особенно симпластотрофобласт, а также децидуальные клетки.
Одним из первых плацента синтезирует хорионический гонадотропин, концентрация которого быстро нарастает на 2-3-й нед беременности, достигая максимума на 8-10-й нед, причем в крови плода она в 10-20 раз выше, чем в крови матери. Гормон стимулирует образование адренокортикотропного гормона (АКТГ) гипофиза, усиливает секрецию кортикостероидов.
Большую роль в развитии беременности играет плацентарный лактоген, который обладает активностью пролактина и лютеотропного гормона гипофиза. Он поддерживает стероидогенез в желтом теле яичника в первые 3 мес беременности, а также принимает участие в метаболизме углеводов и белков. Концентрация его в крови матери прогрессивно нарастает на 3-4-м мес беременности и в дальнейшем продолжает увеличиваться, достигая максимума к 9-му мес. Этот гормон совместно с пролактином гипофиза матери и плода играет определенную роль в продукции легочного сурфактанта и фетоплацен-тарной осморегуляции. Высокая концентрация его обнаруживается в околоплодных водах (в 10-100 раз больше, чем в крови матери).
В хорионе, а также в децидуальной оболочке синтезируются прогестерон и прегнандиол.
Прогестерон (вырабатываемый сначала желтым телом в яичнике, а с 5-6-й нед в плаценте) подавляет сокращения матки, стимулирует ее рост, оказывает иммунодепрессивное действие, подавляя реакцию отторжения плода. Около 3/4 прогестерона в организме матери метаболизируется и трансформируется в эстрогены, а часть выделяется с мочой.
Эстрогены (эстрадиол, эстрон, эстриол) вырабатываются в симпласто-трофобласте ворсин плаценты (хориона) в середине беременности, а к концу
беременности их активность усиливается в 10 раз. Они вызывают гиперплазию и гипертрофию матки.
Кроме того, в плаценте синтезируются меланоцитостимулирующий и адренокортикотропный гормоны, соматостатин и др.
В плаценте содержатся полиамины (спермин, спермидин), влияющие на усиление синтеза РНК в гладких мышечных клетках миометрия, а также на разрушающие их оксидазы. Важную роль играют аминооксидазы (гиста-миназа, моноаминоксидаза), разрушающие биогенные амины - гистамин, серотонин, тирамин. Во время беременности их активность возрастает, что способствует разрушению биогенных аминов и падению концентрации последних в плаценте, миометрии и крови матери.
Во время родов гистамин и серотонин являются наряду с катехоламинами (норадреналин, адреналин) стимуляторами сократительной деятельности гладких мышечных клеток (ГМК) матки, и к концу беременности их концентрация значительно возрастает в связи с резким снижением (в 2 раза) активности аминооксидаз (гистаминаза и др.).
При слабой родовой деятельности отмечается усиление активности аминоокси-даз, например гистаминазы (в 5 раз).
Нормальная плацента не является абсолютным барьером для белков. В частности, фетопротеин в конце 3-го мес беременности проникает в небольшом количестве (около 10 %) из плода в кровь матери, но на этот антиген материнский организм не отвечает отторжением, так как во время беременности уменьшается цитотоксич-ность материнских лимфоцитов.
Плацента препятствует прохождению ряда материнских клеток и цитотоксиче-ских антител к плоду. Главную роль в этом играет фибриноид, покрывающий тро-фобласт при его частичном повреждении. Это предотвращает поступление в межворсинчатое пространство плацентарных и плодовых антигенов, а также ослабляет гуморальную и клеточную «атаку» матери против зародыша.
В заключение отметим основные особенности ранних стадий развития зародыша человека: 1) асинхронный тип полного дробления и образование «светлых» и «темных» бластомеров; 2) раннее обособление и формирование внезародышевых органов; 3) раннее образование амниотического пузырька и отсутствие амниотических складок; 4) наличие в стадии гаструляции двух механизмов - деламинации и иммиграции, в течение которых происходит также развитие провизорных органов; 5) интерстициальный тип имплантации; 6) сильное развитие амниона, хориона, плаценты и слабое развитие желточного мешка и аллантоиса.
21.5. СИСТЕМА МАТЬ-ПЛОД
Система мать-плод возникает в процессе беременности и включает две подсистемы - организм матери и организм плода, а также плаценту, являющуюся связующим звеном между ними.
Взаимодействие между организмом матери и организмом плода обеспечивается прежде всего нейрогуморальными механизмами. При этом в обеих подсистемах различают следующие механизмы: рецепторные, воспринимающие информацию, регуляторные, осуществляющие ее переработку, и исполнительные.
Рецепторные механизмы организма матери расположены в матке в виде чувствительных нервных окончаний, которые первыми воспринимают информацию о состоянии развивающегося плода. В эндометрии находятся хемо-, механо- и терморецепторы, а в кровеносных сосудах - барорецепторы. Рецепторные нервные окончания свободного типа особенно многочисленны в стенках маточной вены и в децидуальной оболочке в области прикрепления плаценты. Раздражение рецепторов матки вызывает изменения интенсивности дыхания, кровяного давления в организме матери, что обеспечивает нормальные условия для развивающегося плода.
Регуляторные механизмы организма матери включают отделы ЦНС (височная доля мозга, гипоталамус, мезэнцефальный отдел ретикулярной формации), а также гипоталамо-эндокринную систему. Важную регуляторную функцию выполняют гормоны: половые, тироксин, кортикостероиды, инсулин и др. Так, во время беременности происходят усиление активности коры надпочечников матери и повышение выработки кортикостероидов, которые участвуют в регуляции метаболизма плода. В плаценте вырабатывается хорионический гонадотропин, стимулирующий образование АКТГ гипофиза, который активизирует деятельность коры надпочечников и усиливает секрецию кортикостероидов.
Регуляторные нейроэндокринные аппараты матери обеспечивают сохранение беременности, необходимый уровень функционирования сердца, сосудов, кроветворных органов, печени и оптимальный уровень обмена веществ, газов в зависимости от потребностей плода.
Рецепторные механизмы организма плода воспринимают сигналы об изменениях организма матери или собственного гомеостаза. Они обнаружены в стенках пупочных артерий и вены, в устьях печеночных вен, в коже и кишечнике плода. Раздражение этих рецепторов приводит к изменению частоты сердцебиения плода, скорости кровотока в его сосудах, влияет на содержание сахара в крови и т. д.
Регуляторные нейрогуморальные механизмы организма плода формируются в процессе развития. Первые двигательные реакции у плода появляются на 2- 3-м мес развития, что свидетельствует о созревании нервных центров. Механизмы, регулирующие газовый гомеостаз, формируются в конце II триместра эмбриогенеза. Начало функционирования центральной эндокринной железы - гипофиза - отмечается на 3-м мес развития. Синтез кортикостероидов в надпочечниках плода начинается со второй половины беременности и увеличивается с его ростом. У плода усилен синтез инсулина, который необходим для обеспечения его роста, связанного с углеводным и энергетическим обменом.
Действие нейрогуморальных регуляторных систем плода направлено на исполнительные механизмы - органы плода, обеспечивающие изменение интенсивности дыхания, сердечно-сосудистой деятельности, мышечной активности и т. п., и на механизмы, определяющие изменение уровня газообмена, обмена веществ, терморегуляции и других функций.
В обеспечении связей в системе мать-плод особо важную роль играет плацента, которая способна не только аккумулировать, но и синтезировать вещества, необходимые для развития плода. Плацента выполняет эндокринные функции, вырабатывая ряд гормонов: прогестерон, эстроген, хориониче-ский гонадотропин (ХГ), плацентарный лактоген и др. Через плаценту между матерью и плодом осуществляются гуморальные и нервные связи.
Существуют также экстраплацентарные гуморальные связи через плодные оболочки и амниотическую жидкость.
Гуморальный канал связи - самый обширный и информативный. Через него происходит поступление кислорода и углекислого газа, белков, углеводов, витаминов, электролитов, гормонов, антител и др. (рис. 21.20). В норме чужеродные вещества не проникают из организма матери через плаценту. Они могут начать проникать лишь в условиях патологии, когда нарушена барьерная функция плаценты. Важным компонентом гуморальных связей являются иммунологические связи, обеспечивающие поддержание иммунного гомеостаза в системе мать-плод.
Несмотря на то, что организмы матери и плода генетически чужеродны по составу белков, иммунологического конфликта обычно не происходит. Это обеспечивается рядом механизмов, среди которых существенное значение имеют следующие: 1) синтезируемые симпластотрофобластом белки, тормозящие иммунный ответ материнского организма; 2) хориональный гонадотропин и плацентарный лактоген, находящиеся в высокой концентрации на поверхности симпластотрофобласта; 3) иммуномаскирую-щее действие гликопротеидов перицеллюлярного фибриноида плаценты, заряженного так же, как и лимфоциты омывающей крови, отрицательно; 4) протеолитические свойства трофобласта также способствуют инактивации чужеродных белков.
В иммунной защите принимают участие и амниотические воды, содержащие антитела, блокирующие антигены А и В, свойственные крови беременной, и не допускают их в кровь плода.
Организмы матери и плода представляют собой динамическую систему гомологичных органов. Поражение какого-либо органа матери ведет к нарушению развития одноименного органа плода. Так, если беременная женщина страдает диабетом, при котором снижена выработка инсулина, то у плода наблюдаются увеличение массы тела и повышение продукции инсулина в островках поджелудочной железы.
В эксперименте на животных установлено, что сыворотка крови животного, у которого удалили часть какого-либо органа, стимулирует пролиферацию в одноименном органе. Однако механизмы этого явления изучены недостаточно.
Нервные связи включают плацентарный и экстраплацентарный каналы: плацентарный - раздражение баро- и хеморецепторов в сосудах плаценты и пуповины, а экстраплацентарный - поступление в ЦНС матери раздражений, связанных с ростом плода и др.
Наличие нервных связей в системе мать-плод подтверждается данными об иннервации плаценты, высоком содержании в ней ацетилхолина, отста-

Рис. 21.20. Транспорт веществ через плацентарный барьер
вании развития плода в денервированном роге матки экспериментальных животных и др.
В процессе формирования системы мать-плод существует ряд критических периодов, наиболее важных для установления взаимодействия между двумя системами, направленных на создание оптимальных условий для развития плода.
21.6. КРИТИЧЕСКИЕ ПЕРИОДЫ РАЗВИТИЯ
В ходе онтогенеза, особенно эмбриогенеза, отмечаются периоды более высокой чувствительности развивающихся половых клеток (в период прогенеза) и зародыша (в период эмбриогенеза). Впервые на это обратил внимание австралийский врач Норман Грегг (1944). Российский эмбриолог П. Г. Светлов (1960) сформулировал теорию критических периодов развития и проверил ее экспериментально. Сущность этой теории
заключается в утверждении общего положения, что каждый этап развития зародыша в целом и его отдельных органов начинается относительно коротким периодом качественно новой перестройки, сопровождающейся детерминацией, пролиферацией и дифференцировкой клеток. В это время эмбрион наиболее восприимчив к повреждающим воздействиям различной природы (рентгеновское облучение, лекарственные средства и др.). Такими периодами в прогенезе являются спермио- и овогенез (мейоз), а в эмбриогенезе - оплодотворение, имплантация (во время которой происходит гаструляция), дифференцировка зародышевых листков и закладка органов, период плацентации (окончательного созревания и формирования плаценты), становление многих функциональных систем, рождение.
Среди развивающихся органов и систем человека особое место принадлежит головному мозгу, который на ранних стадиях выступает в роли первичного организатора дифференцировки окружающих тканевых и органных зачатков (в частности, органов чувств), а позднее отличается интенсивным размножением клеток (примерно 20 000 в минуту), что требует оптимальных условий трофики.
Повреждающими экзогенными факторами в критические периоды могут быть химические вещества, в том числе многие лекарственные, ионизирующее облучение (например, рентгеновское в диагностических дозах), гипоксия, голодание, наркотики, никотин, вирусы и др.
Химические вещества и лекарственные препараты, проникающие через плацентарный барьер, особенно опасны для зародыша в первые 3 мес беременности, так как они не метаболизируются и накапливаются в повышенных концентрациях в его тканях и органах. Наркотики нарушают развитие головного мозга. Голодание, вирусы вызывают пороки развития и даже внутриутробную гибель (табл. 21.2).
Итак, в онтогенезе человека выделяют несколько критических периодов развития: в прогенезе, эмбриогенезе и постнатальной жизни. К ним относятся: 1) развитие половых клеток - овогенез и сперматогенез; 2) оплодотворение; 3) имплантация (7-8-е сут эмбриогенеза); 4) развитие осевых зачатков органов и формирование плаценты (3-8-я нед развития); 5) стадия усиленного роста головного мозга (15-20-я нед); 6) формирование основных функциональных систем организма и дифференцировка полового аппарата (20-24-я нед); 7) рождение; 8) период новорожденности (до 1 года); 9) половое созревание (11-16 лет).
Методы диагностики и меры профилактики аномалий развития человека. С целью выявления аномалий развития человека современная медицина располагает рядом методов (неинвазивных и инвазивных). Так, всем беременным дважды (в 16-24 и 32-36 нед) проводят ультразвуковое исследование, что позволяет обнаружить ряд аномалий развития плода и его органов. На 16-18-й нед беременности с помощью метода определения содержания альфа-фетопротеина в сыворотке крови матери можно выявить пороки развития ЦНС (в случае увеличения его уровня более чем в 2 раза) или хромосомные аномалии, например синдром Дауна - трисомия хромосомы 21 или
Таблица 21.2. Сроки возникновения некоторых аномалий развития эмбрионов и плодов человека

другие трисомии (об этом свидетельствует снижение уровня исследуемого вещества более чем в 2 раза).
Амниоцентез - инвазивный способ исследования, при котором через брюшную стенку матери производят взятие околоплодных вод (обычно на 16-й нед беременности). В дальнейшем производят хромосомный анализ клеток амниотической жидкости и другие исследования.
Используется также визуальный контроль развития плода с помощью лапароскопа, введенного через брюшную стенку матери в полость матки (фетоскопия).
Существуют и другие способы диагностики аномалий развития плода. Однако основной задачей медицинской эмбриологии является предупреждение их развития. С этой целью разрабатываются методы генетического консультирования и подбора супружеских пар.
Методы искусственной инсеминации половыми клетками от заведомо здоровых доноров позволяют избежать наследования ряда неблагоприятных признаков. Развитие генной инженерии дает возможность корригировать локальные повреждения генетического аппарата клетки. Так, существует метод, сущность которого заключается в получении биоптата яичка у
мужчины с генетически обусловленным заболеванием. Внесение в сперма-тогонии нормальной ДНК, а затем трансплантация сперматогоний в предварительно облученное яичко (для уничтожения генетически дефектных половых клеток), последующее размножение трансплантированных спер-матогоний приводит к тому, что вновь образованные сперматозоиды освобождаются от генетически обусловленного дефекта. Следовательно, такие клетки могут дать нормальное потомство при оплодотворении женской половой клетки.
Метод криоконсервации спермы позволяет длительно сохранять оплодотворяющую способность сперматозоидов. Это применяется для сохранения половых клеток мужчин, связанных с опасностью облучения, ранения и др.
Метод искусственного оплодотворения и переноса эмбрионов (экстракорпоральное оплодотворение) применяется для лечения как мужского, так и женского бесплодия. Для получения женских половых клеток используют лапароскопию. Специальной иглой прокалывают оболочку яичника в области расположения пузырчатого фолликула, аспирируют овоцит, который в дальнейшем оплодотворяется спермиями. Последующее культивирование, как правило, до стадии 2-4-8 бластомеров и перенос зародыша в матку или маточную трубу обеспечивает его развитие в условиях материнского организма. При этом возможна трансплантация зародыша в матку «суррогатной» матери.
Совершенствование методов лечения бесплодия и профилактики аномалий развития человека тесно переплетаются с морально-этическими, юридическими, социальными проблемами, решение которых во многом зависит от сложившихся традиций того или иного народа. Это является предметом специального исследования и обсуждения в литературе. В то же время успехи клинической эмбриологии и репродуктологии не могут существенно повлиять на рост народонаселения в силу высокой стоимости лечения и методических трудностей при работе с половыми клетками. Именно поэтому основу деятельности, направленной на оздоровление и численный рост населения, составляет профилактическая работа врача, базирующаяся на знаниях процессов эмбриогенеза. Для рождения здорового потомства немаловажно вести здоровый образ жизни и отказаться от вредных привычек, а также проводить комплекс тех мероприятий, которые находятся в компетенции медицинских, общественных и образовательных учреждений.
Таким образом, в результате изучения эмбриогенеза человека и других позвоночных установлены основные механизмы образования половых клеток и их слияния с возникновением одноклеточной стадии развития - зиготы. Последующее развитие зародыша, имплантация, формирование зародышевых листков и эмбриональных зачатков тканей, внезародышевых органов показывают тесную эволюционную связь и преемственность развития представителей различных классов животного мира. Важно знать, что в развитии зародыша существуют критические периоды, когда резко возрастает риск внутриутробной гибели либо развития по патологическому
пути. Знание основных закономерных процессов эмбриогенеза позволяет решать ряд проблем медицинской эмбриологии (предупреждение аномалий развития плода, лечение бесплодия), осуществлять комплекс мероприятий, предупреждающих гибель плодов и новорожденных.
Контрольные вопросы
1. Тканевый состав детской и материнской частей плаценты.
2. Критические периоды развития человека.
3. Сходство и различия в эмбриогенезе позвоночных и человека.
4. Источники развития тканей провизорных органов.
Гистология, эмбриология, цитология: учебник / Ю. И. Афанасьев, Н. А. Юрина, Е. Ф. Котовский и др.. - 6-е изд., перераб. и доп. - 2012. - 800 с. : ил.
Онтогенезом называют совокупность процессов, протекающих в организме, с момента образования зиготы до смерти.
Его подразделяют на два этапа: эмбриональный ипостэмбриональный.
Эмбриональный период Эмбриональным считают период зародышевого развития с момента образования зиготы до выхода из яйцевых оболочек или рождения, в процессе зародышевого развития эмбрион проходит стадии дробления, гаструляция, первичного органогенеза и дальнейшей дифференцировки органов и тканей. Дробленое . Дроблением называют процесс образования многоклеточного однослойного зародыша - бластулы. Для дробления характерно: 1) деление клеток путем митоза с сохранением диплоидного набора хромосом; 2) очень короткий митотический цикл; 3) бластомеры не дифференцированы, и в них не используется наследственная информация;4) бластомеры не растут и в дальнейшем становятся все меньше; 5) цитоплазма зиготы не перемешивается и не перемещается.
Стадии развития зародыша.
1. Период одноклеточного зародыша, или зиготы, кратковременный, протекающий с момента оплодотворения до начала дробления яйца. 2. Период дробления. В этот период происходит размножениеклеток, Получившиеся при дроблении клетки называют бластомерами. Вначале образуется кучка бластомеров, напоминающая по форме ягоду малины,- морула, затем шаровидная однослойная бластула; стенка бластулы - бластодерма, полость - бластоцеле. 3. Гаструляция. Однослойный зародыш превращается в двухслойный - гаструлу, состоящую из наружного зародышевого листка - эктодермы и внутреннего - энтодермы. У позвоночных уже в ходе гаструляции возникает и третий зародышевый листок - мезодерма. В ходе эволюции у хордовых процесс гаструляции усложнился возникновением осевого комплекса зачатков (закладка нервной системы, осевогоскелетаи мускулатуры) на спинной стороне зародыша. 4. Период обособления основных зачатков органов и тканей и их дальнейшее развитие. Одновременно с этими процессами усиливается объединение частей в единое развивающееся целое. Из эктодермы образуетсяэпителийкожи, нервная система и частичноорганы чувств, из энтодермы - эпителий пищеварительного канала и его железы; из мезодермы - мышцы, эпителий мочеполовой системы и серозных оболочек, из мезенхимы - соединительная, хрящевая и костная ткани, сосудистая система и кровь.
Последствия влияния алкоголя никотина наркотических веществ на зародыш человека.
Систематическое употребление наркотических веществ, к которым относится алкоголь, и даже никотин, вызывает повреждение зародышевых клеток – сперматозоидов и яйцеклеток. Может родиться ребенок с отставанием в длине и массе тела, плохо развивающийся физически, предрасположенный к развитию каких-либо заболеваний. Чем сильнее наркотическое вещество, употребляемое родителями, тем серьезнее могут быть изменения в организме детей. Особенно опасно употребление этих веществ женщинами.
2. Борьба за существование. Предпосылка естественного отбора. Формы борьбы за существование.
Борьба за существование – сложные и многообразные взаимоотношения особей внутри вида, между видами и с неблагоприятными условиями неживой природы. Ч. Дарвин указывает, что несоответствие между возможностью видов к беспредельному размножению и ограниченностью ресурсов – главная причина борьбы за существование. Борьба за существование бывает трех видов:
Внутривидовая
Межвидовая
Борьба с абиотическими факторами
Дробление - это деление зародыша, но это деление несколько отличается от обычного деления митозом: при дроблении отсутствует пресинтетнческая фаза G1, поэтому дочерние (слетки (бластомеры) после митоза не синтезируют белки и не растут - с каждым делением размеры образовавшихся бластомеров становится все меньше и меньше (отсюда и название «‘дробление»). В результате дробления восстанавливается нормальное ядерно-цитоплазматическое отношение (у оплодотворенной яйцеклетки оно очень низкое) и образуется бластула. Тип дробления зависит от количества и распределения желтка в яйцеклетке. Олиголецитальная изолецитальная яйцеклетка человека дробится по типу: полное неравномерное асинхронное. Полное - в дроблении участвуют все участки оплодотворенной яйцеклетки; неравномерное - образовавшиеся бластомеры не одинаковые, не равные: одни крупные и темные, располагаются в центре зародыша их совокупность называется змбриобластом - будущее тело, другие мелкие и светлые, окружают снаружи бластомеры эмбриобласта - их совокупность называется трофобластом - участвует при формировании плаценты; асинхронное = количество бластомеров увеличивается не по геометрической прогреcии (2-> 4-> 8 и т.д., т.е. кратное увеличение числа блаcтомеров). В то же время следует отметить, что первые бластомеры (по крайней мере до 8 первых бластомеров) потенциально не различаются. Доказательством этого утверждения является образование однояйцевых (идентичных) близнецов. Однояйцевые близнецы образуются из одной яйцеклетки в случае когда по каким-то причинам на стадии 2-х или 3-х бластомерного зародыша происходит их разделение и из каждого бластомера развивается самостоятельный отдельный организм. С другой стороны, если в эксперименте взять двухбластомерного зародыша и разрушить, один бластомер, то из второго бластомера может развиться вполне нормальный организм. Или еще; если взять двух эмбрионов на стадии до 8-бласгомерного этапа и их объединить в одну морулу, то можно получить новый организм, наследующий признаки не 2-х а 4-х родителей - такие экспериментально полученные животные называются химерами. Этими экспериментами было доказано, что бластомеры ранних стадий дробления (вплоть до 8-клеточной стадии) практически идентичны и обладают неограниченными потенциями. С помощью метода химер было доказано, что какие бластомеры дадут трофобласт, а какие - эмбриобласт зависит не от особенностей самих бластомеров, а лишь от места, какое они случайно заняли в моруле. Бластомеры расположенные снаружи морулы подвергаются воздействию одних условий, внутренние бластомеры оказываются в других условиях, что и определяет направление их дальнейшей дифференцировки.
Дробление зиготы человека начинается в конце 1-х суток после оплодотворения в дистальной части маточной трубы и заканчивается в 7-е сутки в полости матки. На 2-3-и сутки зародыш находится в маточных трубах и имеет вид плотного узелка - морулы, в центральной части которого находятся крупные темные бластомеры - эмбриоблзст, а по периферии -мелкие светлые бластомеры = трофобласт. В 4-е сутки бластула находится в проксимальной части маточной трубы, т.е. вплотную подходит к полости матки и имеет вид пузырька. Бластомеры трофобласта всасывают секрет маточной трубы и сами секретируют жидкость, поэтому трофобласт растягивается и превращается в пузырек, заполненный жидкостью, а эмбриооласт прикрепляется на одном полюсе к трофобласту снутри. Такая бластула называется эпибластулой (или синонимы; бластоциста, стерробластула). На 5-е сутки бластоциста попадает в полость матки и остается там до 7-х суток, за это время несколько увеличивается в размерах (100 и более бластомеров).
4. Понятие и основные механизмы гаструляции. Типы гаструляции. Строение двухнедельного зародыша. Представление о критических периодах.
Гаструляция - это сложный процесс, где в результате размножения, роста, днфференцировкн н направленного перемещения клеток образуется 3-х листковый зародыш. Гаструляция происходит 7-17 сутки и осуществляется путем деламииацин или расщипления (7-14 сутки) и иммиграции или выселения (14-17 сутки).
В 7-е сутки эмбриобласт расщепляется на 2 слоя: верхний слой - эпибласт или первичная эктодерма (содержит материал будущей эктодермы, мезодермы, хорды и части энтодермы) и нижний слой - гипобласт (будущая энтодерма после присоединения клеточного материала прехордальной пластинки из эпибласта). Почти одновременно с этим происходит выселение клеток из эпи- и гипооласта - внезародышевая мезенхима, которая выстилает внутреннюю поверхность трофобласта. Далее в течение 2-й недели эпибласт и гипобласт начинают прогибаться в противоположных направлениях и превращаются в пузырьки: из эпибласта образуется амниотнческий пузырек, нз гипобласта - желточный пузырек. Эти 2 пузырька окружаются внезародышевой мезенхимой. Соприкасающиеся поверхности амниотнческого и желточного пузырька имеют вид диска (или щитка) и соответственно называются зародышевым эпибластом и зародышевым гипобластом, а вместе - зародышевым щитком. Остальные участки амниогического и желточного пузырька называются внезародышевым эпи- и гипобластом.
В начале 3-й недели (14-17 сутки) происходит иммиграция (выселение) клеток из эпибласта, причем это происходит в 2 фазы: в I фазе идет подготовка к выселению - клеточный материал подлежащий иммиграции перемещается (медленно двигающиеся клетки: с будущего краниального конца к каудальному концу по центру эпибласта, а быстродвигаюшиеся клетки: вначале тоже с краниального полюса к каудальному полюсу, но по краю э:.’нблпста, а затем поворачиваются и идут но центру эпибласта к краниальному концу) и собирается вместе и образует на поверхности эпибласта 3 структуры: прехордальную пластинку, I узелок и I полоску; II фаза - собственно выселение материала этих 3- структур. I узелок выселяется и между имеющимися двумя листками образует первый осевой орган - хорду. Материал прехордальной пластинки выселяется и присоединяется к гипобласту, с этого момента нижним листок будет называться энтодермой.
Клетки 1 полоски выселяется, занимает положение между двумя имеющимися листками и образует средний листок - мезодерму. Оставшаяся часть эпибласта после выселения клеток 3-х структур будет называться эктодермой.
I Эктодерма:
II Мезодерма: -дерматомы -> дерма кожи;
- миотомы -> скелетная мускулатура;
- склеротомы ->
- спланхиотомы ->
III Энтодерма:
IV Мезенхима:
Типы гаструляции
При гаструляции продолжаются изменения, начавшиеся на стадии бластулы, и поэтому разным типам бластул соответствуют и различные типы гаструляции . Переход из бластулы в гаструлу может осуществляться 4-мя основными способами: инвагинацией, иммиграцией, деляминацией и эпиболией.
Инвагинация
или впячивание наблюдается в случае целобластулы. Это наиболее простой способ гаструляции, при котором вегетативная часть впячивается в бластоцель. Вначале появляется небольшое углубление в вегетативном полюсе бластулы. Затем клетки вегетативного полюса все больше и больше впячиваются в полость бластоцеля. В последующем эти клетки доходят до внутренней стороны анимального полюса. Первичная полость, бластоцель, при этом вытесняется и видна только с двух сторон гаструлы в местах изгиба клеток. Зародыш принимает куполообразную форму и становится двухслойным. Его стенка состоит из наружного листка – эктодермы и внутреннего – энтодермы. В результате гаструляции образуется новая полость – гастроцель или полость первичной кишки. Она сообщается с внешней средой с помощью кольцеобразного отверстия – бластопора или первичного рта. Края бластопора называются губами. Различают спинную, брюшную и две боковых губы бластопора.
По последующей судьбе бластопора всех животных разделяют на две большие группы: первично- и вторичноротых. К первичноротым относят животных, у которых бластопор остается постоянным или дефинитивным ртом у взрослой особи (черви, моллюски, членистоногие). У других животных (иглокожие, хордовые) бластопор или превращается в заднепроходное отверстие, или зарастает, а ротовое отверстие возникает заново на переднем конце тела зародыша. Таких животных называют вторичноротыми.
Иммиграция или вселение является наиболее примитивной формой гаструляции. При этом способе происходит перемещение отдельных клеток или группы клеток из бластодермы в бластоцель с образованием энтодермы. Если вселение клеток в бластоцель происходит лишь со стороны одного полюса бластулы, то такая иммиграция называется униполярной, а с различных участков бластулы – мультиполярной. Униполярная иммиграция свойственна некоторым гидроидным полипам, медузам и гидромедузам. В то время, как мультиполярная иммиграция является более редким явлением и наблюдается у некоторых гидромедуз. При иммиграции внутренний зародышевый листок – энтодерма может образовываться сразу в процессе проникновения клеток в полость бластоцеля. В других случаях клетки могут заполнять полость сплошной массой, а затем выстраиваться упорядоченно возле эктодермы и образовывать энтодерму. В последнем случае гастроцель появляется позднее.
Деляминация или расслаивание сводится к расщеплению стенки бластулы. Клетки, которые отделяются внутрь, образуют энтодерму, а наружные – эктодерму. Такой способ гаструляции наблюдается у многих беспозвоночных и высших позвоночных животных.
У некоторых животных в связи с увеличением количества желтка в яйцеклетке и уменьшением полости бластоцеля гаструляция только путем инвагинации становится невозможной. Тогда гаструляция происходит способом эпиболии или обрастания. Этот способ состоит в том, что мелкие анимальные клетки усиленно делятся и обрастают вокруг более крупных вегетативных. Мелкие клетки образуют эктодерму, а клетки вегетативного полюса формируют энтодерму. Такой способ гаструляции наблюдается у круглоротых и земноводных. Периоды наибольшей чувствительности к действию факторов окр среды называются критическими периодами. У человека выделяют 3 основных критич периода в эмбриогенезе:
Имплатация - внедрение эмбриона в слизистую матки (6-7 сутки после оплодотворения)
Плацентация-начало образования плаценты (14-15 сутки)
Роды- выход из матер организма, перестройка функционирования всех систем, изменение способа питания (39-40 неделя). Критич периоды совпадают с переходом от одного периода развития к другому и изменениями условий существования зародыша.
5. Понятие дифференцировки зародышевых листков. Представление об индукции, как фактора, вызывающего дифференцировку. Дифференцировка зародышевых листков у зародыша человека.
Зародышевые листки состоят из клеточных материалов, которые идут на развитие различных органов и тканей. По своему строению клетки различных зародышевых листков отличаются друг от друга; клетки энтодермы всегда крупнее и менее правильной формы, чем эктодермальные. Энтодерма отличается свойствами будущей закладки, имеющей трофическое значение. Эктодерма остается на поверхности и первоначально имеет защитное значение. В отличие от энтодермы она состоит из правильно расположенных клеток более однообразной формы. Гаструляцияприводит к заметному различию между наружным и внутренним листками и зародышевый материал становится неоднородным. Процесс, который приводит к появлению различий в первоначально однородном материале, называется дифференцировкой. Большую роль в дифференцировке клеточного материала играют первичные организаторы или индукторы. Индукторы – это химические вещества, которые выделяются группами клеток и влияют на другие группы клеток, изменяя их путь развития. В результате дифференцировки зародышевых листков образуются различные органы и ткани. При исследовании этих процессов у разных животных было установлено, что судьба каждого зародышевого листка у всех многоклеточных, как правило, одинакова. Индукция (от лат. inductio - побуждение, наведение) в эмбриологии - воздействие одних частей развивающегося зародыша (индукторов) на другие его части (реагирующую систему), осуществляющееся при их контакте и определяющее направление развития реагирующей системы, подобное направлению дифференцировки индуктора (гомотипическая индукция) или отличное от него (гетеротипическая индукция). индукция была открыта в 1901 немецким эмбриологом Х. Шпеманом при изучении образования линзы (хрусталика) глаза из эктодермы у зародышей земноводных. При удалении зачатка глаза линза не возникала. Зачаток глаза, пересаженный на бок зародыша, вызывал образование линзы из эктодермы, которая в норме должна была дифференцироваться в эпидермис кожи. Позже Шпеман обнаружил индуцирующее влияние хордомезодермы на образование из эктодермы гаструлы зачатка центральной нервной системы - нервной пластинки; он назвал это явление первичной эмбриональной индукцей , а индуктор - хордомезодерму -организатором . Дальнейшие исследования с удалением частей развивающегося организма и их культивированием по отдельности или в комбинации и пересадкой в чуждое им место зародыша показали, что явление индукции широко распространено у всех хордовых и многих беспозвоночных животных. Осуществление индукции возможно лишь при условии, что клетки реагирующей системы компетентны к данному воздействию, т. е. способны воспринимать индуцирующий стимул и отвечать на него образованием соответствующих структур. Способность клеток, дифференцирующихся под индуктивным воздействием, самим индуцировать дифференцировку новой группы клеток получило название вторичной индукции .
Во многих случаях установлено, что в процессе индукции не только индуктор влияет на дифференцировку реагирующей системы, но и реагирующая система оказывает на индуктор воздействие, необходимое как для его собственной дифференцировки, так и для осуществления им индуцирующего влияния, т. е. чтоиндукция - взаимодействие групп клеток развивающегося зародыша между собой . Для ряда органогенезов показано, что в процессе индукции из клеток индуктора в клетки реагирующей системы переходят вещества (индуцирующие агенты), которые участвуют в активации синтеза специфических информационных РНК, необходимых для синтеза соответствующих структурных белков в ядрах клеток реагирующей системы.
Действие индукторов, как правило, лишено видовой специфичности. Органоспецифическое действие собств. индукторов может быть в эксперименте заменено действием ряда органов и тканей зародышей старшего возраста и взрослых животных (чужеродные, или гетерогенные, индукторы) или выделенными из них химическими веществами - индуцирующими факторами (напр., из туловищных отделов 9-11-дневных куриных зародышей выделен т. н. вегетализующий фактор - белок с мол. м. ок. 30 000, вызывающий в компетентной эктодерме гаструлы земноводных образование энтодермы и вторично - хорды, мышц и др. производных мезодермы). Действие индукторов может быть имитировано обработкой клеток компетентной ткани более простыми химическими соединениями, например солями натрия и лития, сахарозой, а также некоторыми повреждающими клетки воздействиями; по-видимому, при этом в клетках высвобождаются собств. индуцирующие факторы, находившиеся в них в связанном состоянии. Такую индукцию иногда наз. эвокацией , а индуцирующие стимулы - эвокаторам индукции.
В следующей стадии начинается дифференцировка зародышевых листков в ткани (гистогенез) органов (органогенез) и формирование из органов систем органов (системогенез). При этом следует выделить 20-21 сутки эмбриогенеза - в эти сроки происходят следующие важные процессы:
1 Из всех трех зародышевых листков, но преимущественно нз мезодермы, выселяются клетки, заполняют пространства между тремя зародышевыми листками, т.е. формируется зародышевая мезенхима.
2. Мезодерма дифференцируется на составные части (томы) сомиты, сегментные ножки и спланхнотомы.
3 Трехлистковый плоским зародыш сворачивается «в трубку» - формируется туловище (энтодерма сворачиваясь в трубку превращается к I кишку, окруженную слоем мезенхимы и сплапхнатомами; эктодерма покрывает туловище снаружи).
4. При сворачивании зародыша » в трубку» внезародышевые части организма обособляются от тела зародыша и формируются провизорные органы желточный мешок, гипнотическая оболочка.
Материал зародышевых листков дифференнцируется в ткани, органы и системы органов (более подробно о развитии конкретных органов и систем смотри в лекциях по частной гистологии):
I Эктодерма:
- нервная ткань органов нервной системы;
- эпидермис кожи и его производные (сальные, потовые, молочные железы, ногти, волосы);
- эпителии роговицы и хрусталик глаза, эпителий преддверья ротовой полости и анального отдела прямой кишки;
II Мезодерма: -дерматомы -> дерма кожи;
- миотомы -> скелетная мускулатура;
- склеротомы -> осевой скелет (кости и хрящи позвоночного столба;
- сегментные ножки (нефрототомы) - эпителий мочеполовой системы;
- спланхиотомы -> мезотелий серозных покровов (брюшины, плевры и околосердечной сумки), эпителий гонад (клетки Сертоли яичка и фолликулярные клетки яичников), корковая часть надпочечников, миокард и эпикард;
III Энтодерма:
- эпителий и железы (включая печень и поджелудочную железу) пищеварительном и дыхательной системы;
IV Мезенхима:
ткани внутренней среды (кровь и лимфа, все виды волокнистых соединительных тканей - рыхлая волокнистая соединительная ткань, плотная волокнистая оформленная и неоформленная волокнистая соединительные ткани, соединительные ткани со специальными свойствами, костные и хрящевые ткани) и гладкая мышечная ткань.
Понятие о внезародышевых органах. Внезародышевые органы человека. Образование, строение и значение амниона, желточного мешка и аллантоиса.
Провизорные, или временные, органы образуются в эмбриогенезе ряда представителей позвоночных для обеспечения жизненно важных функций, таких, как дыхание, питание, выделение, движение и др. Недоразвитые органы самого зародыша еще не способны функционировать по назначению, хотя обязательно играют какую-то роль в системе развивающегося целостного организма. Как только зародыш достигает необходимой степени зрелости, когда большинство органов способны выполнять жизненно важные функции, временные органы рассасываются или отбрасываются.
Время образования провизорных органов зависит от того, какие запасы питательных веществ были накоплены в яйцеклетке и в каких условиях среды происходит развитие зародыша. У бесхвостых земноводных, например, благодаря достаточному количеству желтка в яйцеклетке и тому, что развитие идет в воде, зародыш осуществляет газообмен и выделяет продукты диссимиляции непосредственно через оболочки яйца и достигает стадии головастика. На этой стадии образуются провизорные органы дыхания (жабры), пищеварения и движения, приспособленные к водному образу жизни. Перечисленные личиночные органы дают возможность головастику продолжить развитие. По достижении состояния морфофункциональной зрелости органов взрослого типа временные органы исчезают в процессе метаморфоза.
У пресмыкающихся и птиц запасов желтка в яйцеклетке больше, но развитие идет не в воде, а на суше. В связи с этим очень рано возникает потребность в обеспечении дыхания и выделения, а также в защите от высыхания. У них уже в раннем эмбриогенезе, почти параллельно с нейруляцией, начинается формирование провизорных органов, таких, как амнион, хорион и желточный мешок. Чуть позднее формируется аллантоис. У плацентарных млекопитающих эти же провизорные органы образуются еще раньше, поскольку в яйцеклетке очень мало желтка. Развитие таких животных происходит внутриутробно, образование провизорных органов у них совпадает по времени с периодом гаструляции.
Наличие или отсутствие амниона и других провизорных органов лежит в основе деления позвоночных на две группы: Amniota и Anamnia. Эволюционно более древние позвоночные, развивающиеся исключительно в водной среде и представленные такими классами, как Круглоротые, Рыбы и Земноводные, не нуждаются в дополнительных водных и других оболочках зародыша и составляют группу анамний. К группе амниот относят первичноназемных позвоночных, т.е. тех, у кого эмбриональное развитие протекает в наземных условиях.
Это три класса: Пресмыкающиеся, Птицы и Млекопитающие. Они являются высшими позвоночными, так как имеют скоординированные и высокоэффективные системы органов, обеспечивающие им существование в наиболее сложных условиях, каковыми являются условия суши. Эти классы насчитывают большое количество видов, вторично перешедших в водную среду. Таким образом, высшие позвоночные оказались в состоянии освоить все среды обитания. Подобное совершенство было бы невозможным, в том числе и.без внутреннего осеменения и специальных провизорных эмбриональных органов.
В строении и функциях провизорных органов различных амниот много общего. Характеризуя в самом общем виде провизорные органы зародышей высших позвоночных, называемые также зародышевыми оболочками, следует отметить, что все они развиваются из клеточного материала уже сформировавшихся зародышевых листков. Некоторые особенности имеются в развитии зародышевых оболочек плацентарных млекопитающих, о чем будет сказано ниже.
Амнион представляет собой эктодермальный мешок, заключающий зародыша и заполненный амниотической жидкостью. Амниотическая оболочка специализирована для секреции и поглощения амниотической жидкости, омывающей зародыш. Амнион играет первостепенную роль в защите зародыша от высыхания и от механических повреждений, создавая для него наиболее благоприятную и естественную водную среду. Амнион имеет и мезодермальный слой из внезародышевой соматоплевры, который дает начало гладким мышечным волокнам. Сокращения этих мышц вызывают пульсацию амниона, а медленные колебательные движения, сообщаемые при этом зародышу, по-видимому, способствуют тому, что его растущие части не мешают друг другу.
Хорион (сероза) - самая наружная зародышевая оболочка, прилежащая к скорлупе или материнским тканям, возникающая, как и амнион, из эктодермы и соматоплевры. Хорион служит для обмена между зародышем и окружающей средой. У яйцекладущих видов основная его функция - дыхательный газообмен; у млекопитающих он выполняет гораздо более обширные функции, участвуя помимо дыхания в питании, выделении, фильтрации и синтезе веществ, например гормонов.
Желточный мешок имеет энтодермальное происхождение, покрыт висцеральной мезодермой и непосредственно связан с кишечной трубкой зародыша. У зародышей с большим количеством желтка он принимает участие в питании. У птиц, например в спланхноплевре желточного мешка, развивается сосудистая сеть. Желток не проходит через желточный проток, соединяющий мешок с кишкой. Сначала он переводится в растворимую форму под действием пищеварительных ферментов, продуцируемых энтодермальными клетками стенки мешка. Затем попадает в сосуды и с кровью разносится по всему телу зародыша.
У млекопитающих нет запасов желтка и сохранение желточного мешка может быть связано с важными вторичными функциями. Энтодерма желточного мешка служит местом образования первичных половых клеток, мезодерма дает форменные элементы крови зародыша. Кроме того, желточный мешок млекопитающих заполнен жидкостью, отличающейся высокой концентрацией аминокислот и глюкозы, что указывает на возможность обмена белков в желточном мешке.
Судьба желточного мешка у разных животных несколько различна. У птиц к концу периода инкубации остатки желточного мешка уже находятся внутри зародыша, после чего он быстро исчезает и к концу 6-х суток после вылупления полностью рассасывается. У млекопитающих желточный мешок бывает развит по-разному. У хищников он сравнительно большой, с сильно развитой сетью сосудов, а у приматов быстро сморщивается и исчезает без остатка до родов.
Аллантоис развивается несколько позднее других внезародышевых органов. Он представляет собой мешковидный вырост вентральной стенки задней кишки. Следовательно, он образован энтодермой изнутри и спланхноплеврой снаружи. У рептилий и птиц аллантоис быстро дорастает до хориона и выполняет несколько функций. Прежде всего это вместилище для мочевины и мочевой кислоты, которые представляют собой конечные продукты обмена азотсодержащих органических веществ. В аллантоисе хорошо развита сосудистая сеть, благодаря чему вместе с хорионом он участвует в газообмене. При вылуплении наружная часть аллантоиса отбрасывается, а внутренняя - сохраняется в виде мочевого пузыря.
У многих млекопитающих аллантоис тоже хорошо развит и вместе с хорионом образует хориоаллантоисную плаценту. Термин плацента означает тесное наложение или слияние зародышевых оболочек с тканями родительского организма. У приматов и некоторых других млекопитающих энтодермальная часть аллантоиса рудиментарна, а мезодермальные клетки образуют плотный тяж, протягивающийся от клоакального отдела к хориону. По мезодерме аллантоиса к хориону растут сосуды, посредством которых плацента выполняет выделительную, дыхательную и питательную функции.
В процессе эмбрионального развития человека сохраняются общие закономерности развития и стадии, характерные для позвоночных животных. Вместе с тем появляются особенности, отличающие развитие человека от развития других представителей позвоночных; знание этих особенностей необходимо врачу. Процесс внутриутробного развития зародыша человека продолжается в среднем 280 суток (10 лунных месяцев). Эмбриональное развитие человека можно разделить на три периода: начальный (1-я неделя развития), зародышевый (2-8-я неделя развития), плодный (с 9-й недели развития до рождения ребенка). К концу зародышевого периода заканчивается закладка основных эмбриональных зачатков тканей и органов и зародыш приобретает основные черты, характерные для человека. К 9-й неделе развития (начало 3-го месяца) длина зародыша составляет 40 мм, а масса около 5 г. В курсе эмбриологии человека, изучаемом на кафедре гистологии и эмбриологии, основное внимание уделяется особенностям половых клеток человека, оплодотворения и развития человека на ранних стадиях (начальный и зародышевый периоды), когда происходят образование зиготы, дробление, гаструляция, формирование зачатков осевых органов и зародышевых оболочек, гистогенез и органогенез, а также взаимодействия в системе мать - плод. Процессы формирования систем органов у плода подробно рассматриваются в курсе анатомии.
Прогенез
Половые клетки
Мужские половые клетки. Спермин человека образуются в течение всего активного полового периода в больших количествах. Продолжительность развития зрелых сперматозоидов из родоначальных клеток - сперматогоний - составляет около 72 дней. Подробное описание процессов сперматогенеза дается в главе XXII. Сформированный сперматозоид имеет размер около 70 мкм и состоит из головки и хвоста (см. рис. 23). В ядре сперматозоида человека содержится 23 хромосомы, одна из которых является половой (X или V), остальные-аутосомами. Среди спермиев 50% содержат Х-хромосому и 50% - У-хромосому. Показано, что масса Х-хромосомы больше массы У-хромосомы, поэтому спермии, содержащие Х-хромосому, менее подвижны, чем содержащие У-хромосому.
У человека объем эйякулята в норме составляет около 3 мл; в нем содержится в среднем 350 млн. сперматозоидов. Для обеспечения оплодотворения общее количество сперматозоидов в сперме должно быть не менее 150 млн., а концентрация их в 1 мл - не менее 60 млн. В половых путях женщины после копуляцииихчисло уменьшается по направлению от влагалища к дистальному концу маточной трубы. Благодаря высокой подвижности сперматозоиды при оптимальных условиях могут через 30 мин - 1ч достигать полости матки, а через 1 1 / 2 -2 ч находиться в дистальной (ампулярной) части маточной трубы, где происходят встреча с яйцеклеткой и оплодотворение. Спермии сохраняют оплодотворяющую способность до 2 сут.
Женские половые клетки. Образование женских половых клеток (овогенез) совершается в яичниках циклически, при этом в течение овариального цикла каждые 24-28 дней образуется, как правило, один овоцит 1-го порядка (см. гл. XXII). Вышедший из яичника при овуляции овоцит 1-го порядка имеет диаметр около 130 мкм и окружен плотной блестящей зоной, или мембраной, и венцом фолликулярных клеток, число которых достигает 3- 4 тыс. Он подхватывается бахромками маточной трубы (яйцевода) и продвигается по ней. Здесь и заканчивается созревание половой клетки. При этом в результате второго деления созревания образуется овоцит 2-го порядка (яйцеклетка), который утрачивает центриоли и тем самым способность к делению. В ядре яйцеклетки человека содержится 23 хромосомы; одна из них является половой Х-хромосомой.
Яйцеклетка женщины (как и млекопитающих животных) вторично изолецитального типа, содержит небольшое количество желточных зерен, более или менее равномерно расположенных в ооплазме (рис. 32, Л, Б). Свой резерв питательных веществ яйцеклетка человека обычно расходует в течение 12-24 ч после овуляции, а затем погибает, если не будет оплодотворена.
Эмбриогенез
Оплодотворение
Оплодотворение происходит в ампулярной части яйцевода. Оптимальные условия для взаимодействия сперматозоидов с яйцеклеткой обычно создаются в пределах 12 ч после овуляции. При осеменении многочисленные спермии приближаются к яйцеклетке и вступают в контакт с ее оболочкой. Яйцеклетка начинает совершать вращательные движения вокруг своей оси со скоростью 4 вращения в минуту. Эти движения обусловлены влиянием биения жгутиков сперматозоидов и продолжаются около 12 ч. В процессе взаимодействия мужской и женской половых клеток в них происходит ряд изменений. Для спермиев характерны явления капацитации и акросомальная реакция. Капацитация представляет собой процесс активации спермиев, который происходит в яйцеводе под влиянием слизистого секрета его железистых клеток. В механизмах капацитации большое значение принадлежит гормональным факторам, прежде всего прогестерону (гормон желтого тела), активизирующему секрецию железистых клеток яйцеводов. После капацитации следует акросомальная реакция, при которой происходит выделение из сперматозоидов ферментов - гиалуронидазы и трипсина, играющих важную роль в процессе оплодотворения. Гиалуронидаза расщепляет гиалуроновую кислоту, содержащуюся в блестящей зоне. Трипсин расщепляет белки цитолеммы яйцеклетки и клеток лучистого венца. В результате происходят диссоциация и удаление клеток лучистого венца, окружающих яйцеклетку, и растворение блестящей зоны. В яйцеклетке цитолемма в области прикрепления спермия образует приподнимающий бугорок, куда входит один сперматозоид, при этом за счет кортикальной реакции (см. выше) образуется плотная оболочка - оболочка оплодотворения, препятствующая вхождению других спермиев и явлению полиспермии. Ядра женской и мужской половых клеток превращаются в пронуклеусы, сближаются, наступает стадия синкариона. Возникает зигота и к концу 1-х суток после оплодотворения начинается дробление.
Пол будущего ребенка определяется комбинацией половых хромосом в зиготе. Если яйцеклетка оплодотворена сперматозоидом с половой хромосомой X, то в образующемся диплоидном наборе хромосом (у человека их 46) содержатся две Х-хромосомы, характерные для женского организма. При оплодотворении сперматозоидом с половой хромосомой Y в зиготе образуется комбинация половых хромосом XY, характерная для мужского организма. Таким образом, пол ребенка зависит от половых хромосом отца. Так как число образующихся сперматозоидов с Х- и Y-хромосомами одинаково, число новорожденных девочек и мальчиков должно быть равным. Однако в связи с большей чувствительностью эмбрионов мужского пола к повреждающему действию различных факторов число новорожденных мальчиков немного меньше, чем девочек: на 100 мальчиков рождаются 103 девочки.
В медицинской практике выявлены различные виды патологии развития, обусловленные аномальным кариотипом. Причиной подобных аномалий является чаще всего нерасхождение в анафазе половинок половых хромосом в процессе мейоза женских половых клеток. В результате этого в одну клетку попадают две хромосомы и формируется набор половых хромосом XX, а в другую не попадает ни одна. При оплодотворении таких яйцеклеток спермиями с Х или У-половыми хромосомами могут образоваться следующие кариотипы: 1) с 47 хромосомами, из них 3 хромосомы Х (тип XXX) - сверхженский тип, 2) кариотип ОУ (45 хромосом) - нежизнеспособный; 3) кариотип XXY (47 хромосом) - мужской организм с рядом нарушений - уменьшены мужские половые железы, отсутствует сперматогенез, увеличены молочные железы(синдром Клайнфельтера); 4) тип ХО (45 хромосом) -женский организм с рядом изменений - невысокий рост, недоразвитие половых органов (яичника, матки, яйцеводов), отсутствие менструаций и вторичных половых признаков (синдром Тернера).
После образования зиготы начинается зародышевое (эмбриональное) развитие организма. У некоторых организмов (пчел, тлей) эмбриональное развитие может происходить без оплодотворения, т.е. без зиготы. Это явление получило название партеногенез.
Возникающие при этом организмы могут быть гаплоидны (например, самцы пчел) или диплоидны в результате слияния ядер после первого деления или удвоения хромосом без последующего деления клетки.
Из неоплодотворенных яйцеклеток может развиваться новое поколение организмов в естественных условиях или под влиянием различных искусственно созданных воздействий. Такой партеногенез называется искусственным. При помощи него в определенных условиях можно получить даже некоторых позвоночных животных - лягушек, тритонов, индеек.
Отличие развития зародыша у высших растений и животных
Развитие зародыша у высших растений значительно отличается от развития высших животных.
Во-первых, это связано с тем, что у этих растений происходит двойное оплодотворение, после которого начинается развитие как зародыша семени, так и питательного материала - эндосперма.
Во-вторых, у высших животных в период зародышевого развития формируются все ткани и большинство органов, свойственных взрослому, а у зародышей семенных растений образуются только зачатки основных вегетативных органов - корня, стебля и листьев. Полное формирование всех органов, в том числе и таких важнейших, как цветки, происходит в после-зародышевом периоде.
Индивидуальное развитие организма называется онтогенезом.
Первый этап онтогенеза - эмбриональное развитие - в свою очередь, может быть разделен на несколько периодов:
- Дробление;
- гаструляция;
- органогенез — образование тканей и органов зародыша.
Второй этап - формирование тканей и органов зародыша - связан с дальнейшей дифференцировкой клеток. В первую очередь, из эндодермы образуется третий зародышевый листок - мезодерма, который начинает врастать между экто и энтодермой, отделяя их друг от друга.
Затем у зародышей позвоночных животных начинается формирование нервной трубки и хорды . Нервная трубка образуется на будущей спинной стороне зародыша путем впячивания эктодермы в виде желобка. Края желобка о дальнейшем срастаются, и он превращается в трубку, которая погружается под эктодерму. Нервная трубка является зачатком спинного мозга .
В это же время под нервной трубкой из энтодермы образуется тяж клеток, который впоследствии формирует хорду.
Дальнейшая дифференцировка приводит к формированию из энтодермы эпителия кишок, пищеварительных желез, а также легких.
Из мезодермы образуются кровеносная, выделительная система, скелет, мышцы. Из эктодермы кроме нервной трубки образуются органы чувств, покровный эпителий и придатки кожи.
Взаимодействие и взаимовлияние частей развивающегося зародыша
В процессе эмбрионального развития одни ткани или органы зародыша могут оказывать влияние на развитие других, находящихся рядом. Это влияние осуществляется путем сложных биохимических воздействий одних частей зародыша на другие. Такое влияние, определяющее направление развития, называется индукцией .
Если пересадить зачаток глаза, взятого у одного зародыша, под эктодерму другого зародыша, из последней развивается хрусталик дополнительного глаза. В этом случае пересаженный участок индуцировал дифференцировку клеток эктодермы в зачаток хрусталика. Индукцию можно наблюдать при пересадке спинного края бластопора одного зародыша на стадии гаструлы другому на той же стадии.
Этот участок бластопора является индуктором образования осевых органов зародыша - хорды и нервной трубки. В результате у зародыша образуются два комплекса осевых органов - один под влиянием собственного края бластопора, другой - под влиянием пересаженного. В некоторых случаях при этом удается получить два сращенных вместе зародыша.
Эмбриональный период заканчивается выходом зародыша из яйцевых оболочек. Далее начинается постэмбриональное развитие, которое характеризуется переходом организма к самостоятельному питанию и активному движению.