Т-клетки это фактически приобретенный иммунитет, способный защитить от цитотоксического повреждающего воздействия на организм. Чужеродные клетки-агрессоры, попадая в организм вносят “хаос”, который внешне проявляется в симптомах заболеваний.
Клетки-агрессоры повреждают по ходу своей деятельности в организме все, что могут, действуя в своих интересах. А задача иммунной системы найти и уничтожить все чуждые элементы.
Специфическая защита организма от биологической агрессии (чужеродных молекул, клеток, токсинов, бактерий, вирусов, грибов и т.д.) осуществляется при помощи двух механизмов:
- продуцирования специфических антител в ответ на чужеродные антигены (потенциально опасных для организма веществ);
- выработки клеточных факторов приобретённого иммунитета (Т-клетки).
При попадании в организм человека «клетки-агрессора», иммунная система распознаёт чужеродные и собственные измененные макромолекулы (антигены) и удаляет их из организма. Также при первичном контакте с новыми антигенами происходит их запоминание, что способствует их более быстрому удалению, в случае вторичного попадания в организм.
Процесс запоминания (презентация) происходит благодаря антиген-распознающим рецепторам клеток и работе антиген представляющих молекул (МНС молекул- комплексов гистосовместимости).
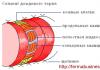
Что такое Т-клетки иммунной системы, и какие функции они выполняют
Функционирование иммунной системы обуславливается работой
. Это клетки иммунной системы, являющиеся  разновидностью лейкоцитов и способствующие формированию приобретённого иммунитета. Среди них выделяют:
разновидностью лейкоцитов и способствующие формированию приобретённого иммунитета. Среди них выделяют:
- В-клетки (распознающие «агрессора» и вырабатывающие к нему антитела);
- Т-клетки (выполняющие функцию регулятора клеточного иммунитета);
- NК- клетки (разрушающие отмеченные антителами чужеродные структуры).
Однако, помимо регуляции иммунного ответа, Т-лимфоциты способны выполнять эффекторную функцию, разрушая опухолевые, мутировавшие и чужеродные клетки, участвовать в формировании иммунологической памяти, распознавать антигены и индуцировать иммунные реакции.
Справочно. Важной особенностью T-клеток является их способность реагировать только на презентированные антигены. На одном T-лимфоците находится только один рецептор к одному конкретному антигену. Это обеспечивает отсутствие реакции T-клеток на собственные аутоантигены организма.
Разнообразие функций Т-лимфоцитов обусловлено наличием в них субпопуляций, представленных Т-хелперами, Т-киллерами и Т-супрессорами.
Субпопуляция клеток, их стадия дифференцирования (развития), степень зрелости и т.д. определяется при помощи специальных кластеров дифференцировки, обозначаемых как СD. Наиболее значимыми являются СD3, СD4 и СD8:
- СD3 находится на всех зрелых T-лимфоцитах, он способствует передаче сигнала от рецептора к цитоплазме. Это важный маркер функционирования лимфоцитов.
- СD8 – это маркер цитотоксических T-клеток.
- СD4 является маркером T-хелперов и рецептором к ВИЧ (вирус иммунодефицита человека)
Читайте также по теме
Гемотрансфузионные осложнения при переливании крови
Т-хелперы
Около половины Т-лимфоцитов имеют антиген CD4, то есть являются Т-хелперами. Это помощники, стимулирующие процесс секреции антител В-лимфоцитами, стимулируют работу моноцитов, тучных клеток и предшественников Т-киллеров к «включению» в иммунную реакцию.Справочно. Функция хелперов осуществляется за счёт синтеза цитокинов (информационных молекул, регулирующих взаимодействие между клетками).
В зависимости от продуцируемого цитокина, их разделяют на:
- T-хелперные клетки 1-го класса (продуцируют интерлейкин-2 и гамма-интерферон, обеспечивая гуморальный иммунный ответ на вирусы, бактерии, опухоли и трансплантанты).
- T-хелперные клетки 2-го класса (секретируют интерлейкины-4,-5,-10,-13 и отвечают за образование IgЕ, а также иммунного ответа, направленного на внеклеточные бактерии).
Т-хелперы 1-го и 2-го типа всегда взаимодействуют антагонистически, то есть повышенная активность первого типа угнетает функцию второго типа и наоборот.
Работа хелперов обеспечивает взаимодействие между всеми клетками иммунитета, определяя какой тип иммунного ответа будет преобладать (клеточный либо гуморальный).
Важно. Нарушение работы клеток-помощников, а именно недостаточность их функции, наблюдается у больных с приобретённым иммунодефицитом. Т-хелперы являются основной мишенью ВИЧ. В результате их гибели нарушается иммунная реакция организма на стимуляцию антигенов, что приводит к развитию тяжёлых инфекций, росту онкологических новообразований и летальному исходу.
 Это так называемые T-эффекторы (цитотоксические клетки) или клетки убийцы. Такое название обусловлено их способностью уничтожать клетки-мишени. Осуществляя лизирование (ли́зис (от греч. λύσις - разделение) - растворение клеток и их систем) мишеней, переносящих чужеродный антиген или мутировавший аутоантиген (трансплантанты, опухолевые клетки), они обеспечивают реакции противоопухолевой защиты, трансплантационного и противовирусного иммунитета, а также аутоиммунные реакции.
Это так называемые T-эффекторы (цитотоксические клетки) или клетки убийцы. Такое название обусловлено их способностью уничтожать клетки-мишени. Осуществляя лизирование (ли́зис (от греч. λύσις - разделение) - растворение клеток и их систем) мишеней, переносящих чужеродный антиген или мутировавший аутоантиген (трансплантанты, опухолевые клетки), они обеспечивают реакции противоопухолевой защиты, трансплантационного и противовирусного иммунитета, а также аутоиммунные реакции.
Т-киллеры при помощи собственных МНС-молекул распознают чужеродный антиген. Связываясь с ним на поверхности клетки, они продуцируют перфорин (цитотоксический белок).
После лизирования клетки «агрессора» Т-киллеры остаются жизнеспособными и продолжают циркулировать в крови, разрушая чужеродные антигены.
Т-киллеры составляют до 25-ти процентов от всех Т-лимфоциотов.
Справочно. Помимо обеспечения реакций нормального иммунного ответа, Т-эффекторы могут участвовать в реакциях антителозависимой клеточной цитотоксичности, способствуя развитию гиперчувствительности второго типа (цитотоксической).
Это может проявляться лекарственными аллергиями и различными аутоиммунными заболеваниями (системные заболевания соединительной ткани, гемолитическая анемия аутоиммунного характера, злокачественная миастения, аутоиммунные тиреоидиты, и т.д.).
Подобным механизмом действия обладают некоторые лекарственные средства, способные запускать процессы некроза опухолевых клеток.
Важно. Препараты с цитотоксическим действием используют в химиотерапии онкологических заболеваний.
Например, к таким медикаментам относится Хлорбутин. Это средство применяют для лечения хронического лимфолейкоза, лимфогранулематоза и рака яичников.
Лимфоциты, как и другие клетки иммунной системы, являются производными полипотентной стволовой клетки костного мозга. В результате пролиферации и дифференцировки стволовых клеток формируются две основные группы лимфоцитов, именуемые В- и Т- лимфоцитами, которые морфологически не отличимы друг от друга (схема 13.1).
Морфологически лимфоцит – клетка шаровидной формы с большим ядром и узким слоем базофильной цитоплазмы. В процессе дифференцировки последовательно формируются большие, средние и малые лимфоциты. В лимфе и периферической крови большинство составляют наиболее зрелые малые лимфоциты, которые обладают амебовидной подвижностью. Они постоянно перемещаются с током лимфы или крови, накапливаясь в лимфоидных органах и тканях, где осуществляются иммунологические реакции.
Две основные популяции лимфоцитов Т- и В-клетки при световой микроскопии не различаются, но четко дифференцируются по поверхностным структурам и функциональным свойствам. Их сравнительные характеристики представлены в табл. 13.2.
Основные функциональные отличия Т - и В -лимфоцитов состоят в том, что В -лимфоциты осуществляют гуморальный иммунный ответ, а Т - лимфоциты – клеточный, а также участвуют в регуляции обеих форм иммунного ответа; при этом Т -система по отношению к В -системе является регулирующей.
Т -лимфоциты получили обозначение потому, что созревают и дифференцируются в тимусе. Они составляют около 80% всех лимфоцитов крови и лимфоузлов, содержатся во всех тканях организма.
Они осуществляют две основные функции – регуляторную и эффекторную.
Регуляторные клетки обеспечивают развитие иммунного ответа другими клетками, регулируют его дальнейшее течение.
Эффекторные Т -лимфоциты осуществляют эффект иммунологической реакции чаще всего в форме цитолиза клеточных структур, к антигенам которых возникла иммунологическая реакция.
Все Т-лимфоциты обладают поверхностными молекулами CD2, определяющими их адгезивные свойства и молекулами CD3, являющимися рецепторами для антигенов. В тимусе Т- лимфоциты дифференцируются на две субпопуляции, содержащие антигены CD4 либо CD8.
Лимфоциты CD4 обладают свойствами клеток – помощников – хелперов (Тх), лимфоциты CD8 – цитотоксическими свойствами, а также супрессорным эффектом, заключающимся в их способности подавлять активность других клеток иммунной системы.
В ответ на антигенный стимул Т-лимфоциты трансформируются в иммунобласты – крупные делящиеся клетки с пиронинофильной цитоплазмой, содержащей многочисленные рибосомы и полирибосомы. Иммунобласты Т-клеток синтезируют и экскретируют в окружающую среду растворимые факторы (лимфокины), являющиеся медиаторами иммунитета.
Т-иммунобласты неоднородны по своему функциональному участию в регуляции иммунного ответа. Они дифференцируются на следующие популяции Т -лимфоцитов:
1. Т -киллеры (tokill – убивать) или син. Т -эффекторы – они обладают специфической цитотоксической активностью в отношении клеток – мишений без участия антител и комплемента. Клетка – киллер оказывает действие в результате прямого контакта с антигенными детерминантами клетки – мишени. Т- эффекторы ответственны за клеточный иммунитет в различных его проявлениях: разрушают опухолевые клетки, трансплантированные клетки, мутировавшие клетки собственного организма, участвуют в гиперчувствительности замедленного типа. Это цитоцидные клетки, разрушающие клетки-мишени при непосредственном контакте за счет выделяемых ферментов-токсинов или в результате активации в клетках-мишенях лизосомальных ферментов.
2. Т -хелперы (tohelp –помогать) относятся к регулирующим клеткам. Получив от макрофагов информацию об антигене, Т-хелперы с помощью иммуноцитокинов передают сигнал, усиливающий пролифирацию Т- и В -лимфоцитов нужных клонов, превращая их в активированные Т-эффекторы или, взаимодействуя с В 2 -лимфоцитами, стимулируют их трансформацию в плазмоциты, которые синтезируют антитела.
3. Т -супрессоры (suppression – подавление) тоже относятся к регуляторам иммунного ответа. Они являются антагонистами Т-хелперов, т. е. блокируют Т- хелперы, тормозят пролиферацию иммунокомпетентных В- клеток, способствуют развитию толерантности. Действие Т- супрессоров позволяет ограничить силу иммунного ответа биологической потребностью, достаточной для восстановления гомеостаза, предотвратить избыточную продукцию иммуноглобулинов. Гиперфункция Т- супрессоров сопровождается угнетение иммунного ответа, вплоть до полного его подавления. Недостаточность Т- супрессоров ведет к развитию аутоиммунных и других вредных для организма реакций.
4. Т -усилители, или Т - амплифайеры (amplifier – усилитель) выполняют функцию помощников в иммунном ответе клеточного типа, а именно: усиливают действие тех или иных субпопуляций Т-лимфоцитов.
5. Т -дифференцирующие клетки (difference – различие) изменяют дифференцировку стволовых клеток гемопоэза в миелоидном или лимфоидном направлениях.
6. Т -лимфоциты иммунологической памяти (immunememori) – стимулированные антигеном Т - лимфоциты, способные сохранять и передавать другим клеткам информацию об этом антигене. При повторном попадании в организм антигена клетки памяти обеспечивают его иммунное распознавание и ответ по вторичному типу.
К цитотоксическим лимфоцитам (Т- киллеры) по происхождению и функциям близки естественные киллеры (ЕК), которые имеют общих предков – предшественников с Т-лимфоцитами. Однако ЕК не попадают в тимус и не подвергаются дифференцировке и селекции. Эти лимфоциты не имеют рецепторов для антигенов и поэтому не участвуют в специфических реакциях приобретенного иммунитета. ЕК относятся к системе естественного иммунитета и разрушают в организме любые клетки, зараженные вирусами, а также опухолевые клетки. В отличие от цитотоксических Т- лимфоцитов, формирующихся и проявляющих свое действие в организме только после антигенной стимуляции, ЕК всегда готовы к контакту с мишенями и цитотоксическому действию. Механизмы их цитотоксического действия сходны с действием Т- киллеров (т. е. за счет образования активных субстратов). Маркерами ЕК человека служат поверхностные антигены СD 56, СD 16 (и СD 2). Сами ЕК продуцируют цитокины, активирующие другие клетки иммунной системы, повышая общий уровень защитных реакций.
В -лимфоциты составляют вторую основную популяцию лимфоцитов. Эти клетки составляют 10 – 15 % лимфоцитов крови, 20 – 25 % клеток лимфоузлов.
В-лимфоциты выполняют в организме две роли: обеспечивают продукцию антител и участвуют в представлении антигенов В-лимфоцитам.
В-лимфоциты обладают поверхностными рецепторами для антигенов, представляющих собой молекулы иммуноглобулинов, чаще всего классов D и М, фиксированных на их наружной мембране. На поверхности одного
В-лимфоцита находится 200-500 тыс. молекул одинаковой специфичности. Отделившиеся от В-лимфоцита иммуноглобулиновые рецепторы циркулируют в организме как свободные антитела.
В-лимфоцит происходит от стволовой кроветворной клетки, проходит созревание в костном мозге, где на его поверхности формируются иммуноглобулиновые рецепторы для антигенов. На каждом лимфоците формируются рецепторы только для одного антигена. Созревающий лимфоцит покидает костный мозг и становится антиген-реактивной клеткой, т. е. клеткой, способной к взаимодействию с одним из многочисленных антигенов, существующих в природе. В отличие от Т-лимфоцитов, которые могут взаимодействовать с антигеном только после его представления антиген-представляющей клеткой, В-лимфоциты вступают в контакт с антигеном напрямую, без посредников. Контакт с антигеном может служить стимулом для пролиферации и дифференцировки В-лимфоцитов.
В-лимфоциты последовательно превращаются в иммуноциты, плазмобласты и плазмоциты.
Плазмоциты – основные клетки, которые синтезируют и экскретируют антитела. Плазматическая клетка относится к короткоживущим клеткам. Плазмоциты не имеют на наружной мембране рецепторов для антигена. Они – конечный продукт дифференцировки В-лимфоцитов. Интенсивность синтеза иммуноглобулинов одной плазматической клеткой достигает 1 млн молекул в час. После завершения фазы активной продукции антител плазмоциты прекращают свое существование.
В популяции В -лимфоцитов различают несколько субпопуляций:
1. В 1 -лимфоциты – предшественники плазмоцитов, синтезирующих антитела без взаимодействия с Т- хелперами. Есть тимуснезависимые антигены (бактериальные полисахариды, полимеризованный флагелин, леван и др.), которые способны без Т-лимфоцитов реагировать, т. е. фиксироваться на рецепторах В-клеток. Эти антигены стимулируют синтез только Ig М.
2. В 2 –лимфоциты, превращаются после антигенной стимуляции в плазмоциты с помощью Т-хелперов, ответственны за гуморальный ответ на тимусзависимые антигены, сопровождающийся синтезом иммуноглобулинов всех классов.
3. В 3 -лимфоциты (В -киллеры) оказывают цитотоксическое действие на клетки-мишени, покрытые антителами, без участия комплемента. Допускается, что В-киллеры – производные «нулевых» лимфоцитов – лимфоцитов без отличительных признаков Т- и В- клеток. Тот факт, что они встречаются среди лимфоцитов костного мозга в 50 % случаев, а среди лимфоцитов крови в 5% случаев, позволяет предположить, что это незрелые формы лимфоцитов, хотя и обладающие цитотоксической активностью.
4. В -супрессоры тормозят пролиферацию и трансформацию Т-клеток, стимулированных антигеном. Супрессорное действие В-клеток, как и Т-клеток, осуществляется при непосредственном контакте с иммунокомпетентными клетками и опосредованно через медиаторы.
5. В -лимфоциты памяти формируются в ходе иммунного ответа на антиген, составляют около 1 % всех В-лимфоцитов, отличаются долголетием и способностью быстро отвечать на повторное поступление антигена. В- лимфоциты памяти не имеют морфологических отличий от других В -лимфоцитов, но обладают активным геном (bcl-2). В-клетки памяти рециркулируют между кровью, лимфой и лимфоидными органами, но более всего накапливаются в периферических лимфоидных органах. Они сохраняют информацию об антигене, способны передавать её другим клеткам, обеспечивают при повторном попадании антигена синтез Ig по вторичному признаку.
Макрофаги – это антигенпредставляющие клетки (АПК), т.к. они обладают антигенами МНС II класса и способностью сорбировать на своей поверхности чужеродный антиген. Макрофаги, дендритные клетки и
В-лимфоциты называют профессиональными АПК, т. к. они более мобильны, активны и выполняют основной объем функций представления антигенов. АПК имеет на наружной мембране до 2 . 10 5 молекул МНС II класса. Для активации одного Т-лимфоцита достаточно 200 – 300 таких молекул, находящихся в комплексе с антигеном.
Макрофаги развиваются из миелопоэтической стволовой клетки костного мозга, проходя этапы: промоноцит – циркулирующий моноцит – тканевой макрофаг.
Моноциты, составляющие около 5 % лейкоцитов крови, находятся в циркуляции около 1 суток, а затем поступают в ткани, формируя популяцию тканевых макрофагов, количество которых в 25 раз больше, чем моноцитов. К ним относятся купферовские клетки печени, микроглия центральной нервной системы, остеокласты костной ткани, макрофаги легочных альвеол, кожи и других тканей. Много макрофагов во всех органах иммунной системы.
Тканевые макрофаги – клетки с округлым или почковидным ядром имеют диаметр 40 – 50 мкм. Цитоплазма содержит лизосомы с набором гидролитических ферментов, обеспечивающих переваривание любых органических веществ и выделение бактерицидного аниона кислорода.
Макрофаги функционируют как фагоциты.
Участие макрофага в иммунном ответе состоит в том, что эта клетка фагоцитирует антиген-содержащие частицы, дезинтегрирует их, превращая белки в антигенные пептидные фрагменты. Последние в комплексе с собственными антигенами МНС II класса макрофаг передает Т-лимфоциту при прямом контакте с ним.
При этом макрофаг продуцирует лимфокин ИЛ – 1 , который вызывает пролиферацию лимфоцитов, вступивших в контакт с антигеном, что обеспечивает формирование клона этих клеток, осуществляющих развитие иммунологической реакции на антиген.
Дендритные клетки составляют вторую группу АПК. Они близки к макрофагам, но не обладают фагоцитирующими свойствами. Это способствует сохранности поглощенных антигенов. Дендритные клетки содержатся в крови, лимфе и во всех других тканях. Дендритные клетки эпителиальных тканей называют клетками Лангерганса, в лимфоузлах и селезенке они составляют около 1 % всех клеток. Эти отростчатые мононуклеарные клетки в разных тканях имеют неодинаковую форму и даже названия, однако все они обладают молекулами МНС II класса и способностью фиксировать антигены с формированием комплекса антиген-продукт МНС, представляемого Т-лимфоцитам.
Дендритные клетки более активны, чем макрофаги и В- клетки в индукции первичного иммунного ответа: в отличие от других АПК дендритные клетки могут представлять антиген покоящимся Т – лимфоцитам. Захват антигена дендритными клетками чаще всего происходит вне лимфоидных органов. После этого они мигрируют в лимфоидные образования, где происходит их контакт с Т-лимфоцитами и развитие дальнейших событий иммунного ответа. МНС II класса – молекула, представляющая антиген CD4 Т-лимфоциту хелперу, а МНС I класса – молекула, представляющая антиген CD8 Т-лимфоциту киллеру. Поэтому дендритные клетки являются также инициаторами цитотоксических реакций.
В -лимфоциты как АПК в отличие от других АПК вступают в контакт с антигеном через свои специфические рецепторы. Следовательно, в представлении антигена участвуют не все В - лимфоциты, а только те, которые обладают рецепторами к данному антигену. Вследствие этого для индукции иммунного ответа требуется в 10 тыс. раз меньше антигена, чем при его представлении другими АПК. Процесс присоединения антигена к В -лимфоциту длится несколько минут, после чего антиген подвергается эндоцитозу. Далее В -лимфоцит вступает в прямой контакт с Т-клеткой и служит сигналом для ее активации.
Клетки антиген - неспецифической резистентности
В осуществлении иммунной защиты организма принимают участие клетки, которые не распознают антигены как лимфоциты и не представляют их лимфоцитам, как АПК.
Это клетки группы гранулоцитов, которые обладают способностью отличать клетки собственного организма от чужеродных, подвергать последние фагоцитозу и индуцировать воспалительные реакции.
Такие же свойства присущи моноцитам, макрофагам и их производным – клеткам, участвующим как в реакциях естественного иммунитета, так и в индукции специфического иммунного ответа в качестве АПК.
Нейтрофильные, базофильные, эозинофильные лейкоциты, а также макрофаги продуцитруют цитокины, регулирующие активность лимфоцитов и сами находятся под их контролем. Эозинофилы обеспечивают наиболее эффективный фагоцитоз гельминтов. Базофильные лейкоциты и тучные клетки содержат в цитоплазме до 100 – 500 гранул, содержащих гистамин, гепарин, серотонин и другие медиаторы, которые выходя из клетки оказывают повреждающее действие как на микроорганизмы, так и на собственные окружающие клетки, способствуя развитию анафилактической реакции.
Кровяные пластинки, или тромбоциты, относятся к системе свертывания крови и играют существенную роль в воспалительных реакциях, регулируют циркуляцию клеток, фиксацию иммунных комплексов в тканях. Тромбоциты содержат медиаторы аллергических реакций, прямо способствующие развитию аллергического воспаления.
Несмотря на большое разнообразие, система клеток и органов иммунной системы функционирует как единое целое на основе единства и функционального программирования всех ее элементов, межклеточной кооперации, механизмов обратной связи, а также антигеннеспецифической регуляции всей системы цитокинами, гормональными и метаболическими механизмами.
Для полного иммунного ответа на большинсво антигенов необходимо взаимодействие макрофагов с Т - и В - лимфоцитами.
Основные иммунологические феномены включают:
1) гуморальные факторы (антителообразование); 2)клеточные факторы.
Лимфоциты - это особые клетки в организме живого существа. Именно они отвечают за его защиту от внешних раздражителей, инфекций, вирусов. Но само понятие "лимфоциты" - довольно обширное и общее. Внутри себя эти клетки будут разделяться еще на несколько групп. В статье мы подробно познакомимся с одной из них - Т-лимфоцитами. Функции, разновидности клеток, нормальные их показатели, отклонения от нормы в крови человека - все эти темы будут рассмотрены далее.
Происхождение клеток
Где образуются клетки Т-лимфоциты? Хоть основное место их "жительства" - кровеносное русло (лимфоциты также обитают и в иных тканях), образуются они далеко не там. Место их "рождения" - костный красный мозг. Он известен как кроветворная ткань организма. То есть, помимо лимфоцитов, тут также будут образовываться эритроциты, белые кровяные клетки (нейтрофилы, лейкоциты, моноциты).
Строение лимфоцитов
"Анатомические" особенности следующие:
- Крупное ядро круглой или овальной формы.
- В цитоплазме (содержимом самой клетки) будет отсутствовать зернистость.
- Если цитоплазмы в клетке мало, она называется узкоплазменной, если много - широкоплазменной.
По своему строению лимфоциты, населяющие кровь, будут немного отличаться от своих собратьев, поселившихся в иных тканях. И это нормально. Более того, клетки, "живущие" в одном месте, тоже будут иметь между собой некоторые внешние различия.
Виды лимфоцитов
Кроме типов Т-лимфоцитов, существуют различные группировки этих клеток вообще. Давайте рассмотрим их.
Первая классификация - по размеру:
- Малые.
- Большие.
Вторая классификация - по выполняемым функциям:
- В-лимфоциты. Могут распознавать инородные частицы и вырабатывать против них убийственные антитела. Иными словами, ответственны за гуморальный иммунитет.
- Т-лимфоциты. Основная функция - ответственность за клеточный иммунитет. Вступают в контакт с чужеродными телами и уничтожают их.
- NK-клетки. Натуральные киллеры, что могут распознать раковые, дефектные клетки и уничтожить их. Отвечают за сохранение клеточного нормального состава всего организма.

Разновидности Т-лимфоцитов
Данная группа лимфоцитов внутри себя будет разделяться еще на несколько типов:
- Т-киллеры.
- Т-хелперы.
- Т-супрессоры.
- Т-клетки памяти.
- Ампликаторы-лимфоциты.

Т-киллеры: что за тип
Это самые известные представители группы Т-лимфоцитов. Их главная задача - разрушение неполноценных, дефектных клеток организма. Иное название группы - цитотоксические Т-лимфоциты. Иными словами, отвечающие за устранение клеток ("цито"), которые обладают токсическим воздействием на весь организм.
Главная функция Т-киллеров - иммунный надзор. Клетки агрессивно воздействуют на чужеродный белок. Именно эта полезная функция бывает вредной при пересадке человеку органов. Т-киллеры стремятся поскорее уничтожить "чужака", не понимая, что именно он способен спасти организм. Поэтому пациент какое-то время после трансплантации органа принимает медикаменты, что угнетают иммунную систему. Препараты снижают процент Т-киллеров в крови, нарушают их взаимодействие. Благодаря этому пересаженный орган приживается, а пациенту не грозят осложнения и летальный исход.
Механизм воздействия данного вида лимфоцитов на чужеродный элемент очень интересен. Фагоциты, например, агрессивно "нападают" на "чужака" для его последующего пожирания и переваривания. Т-киллеры на их фоне - "благородные убийцы". Они прикасаются к объекту своими отростками, затем разрывают контакт и удаляются. Только после такого "смертельного поцелуя" инородный микроорганизм погибает. Почему же?
Во время прикосновения Т-киллеры оставляют на поверхности организма частичку своей мембраны. Она обладает свойствами, которые позволяют ей разъедать поверхность объекта нападения - вплоть до образования сквозных дыр. Через эти отверстия из микроорганизма уходят ионы калия, а их место занимает вода и ионы натрия. Клеточный барьер нарушается, между внутренней и внешней средой уже нет границы. Микроорганизм раздувает поступившая в него вода, белки цитоплазмы и органеллы разрушаются. Остатки "чужака" после этого пожирают фагоциты.

Хелперы
Главная функция этих клеток Т-лимфоцитов - помощь. Отсюда и их название, произошедшее от английского слова, переводящегося так же.
Но кому или чему приходят на выручку данные Т-лимфоциты? Они призваны индуцировать и стимулировать иммунный ответ. Именно под влиянием Т-хелперов будут активировать свою работу Т-киллеры, с которыми мы уже успели познакомиться.
Хелперы станут передавать данные о присутствии в организме чужеродного белка. А это ценная информация для В-лимфоцитов - они, в свою очередь, начинают выделять против него определенные защитные антитела.
Также Т-хелперы стимулируют работу еще одной разновидности "охранных" клеток - фагоцитов. В частности, они плотно взаимодействуют с моноцитами.
Супрессоры
Этот термин сам по себе означает "подавление". Отсюда нам становится понятной функция Т-супрессоров. Хелперы в нашем организме будут активировать защитную, иммунную функцию, а данные Т-лимфоциты, напротив, будут ее подавлять.
Не надо думать, что это оказывает какое-то негативное влияние на систему. Т-супрессоры отвечают за регуляцию иммунного ответа. Ведь где-то нужно реагировать на определенный раздражитель сдержанно и умеренно, а где-то - аккумулировать против него все имеющиеся силы.

Амплификаторы
Перейдем теперь к функциям Т-лимфоцитов данной группы. После того как в организм проникает тот или иной агрессор, в крови и тканях живого существа сразу же повышается содержание лимфоцитов. К примеру, буквально за несколько часов их объем может увеличиться в два раза!
Какова же причина такого быстрого роста армии клеток-защитников? Может быть, дело в том, что в организме где-то до поры до времени они "припрятаны" про запас?
Это действительно так. В тимусе и селезенке обитает какая-то масса зрелых полноценных лимфоцитов. Только до какого-то момента эти клетки "не определяются" со своим предназначением, функцией. Они и будут называться амплификаторами. При надобности данные клетки превращаются в ту или иную разновидность Т-лимфоцитов.
Клетки памяти
Опыт, как известно, - главное оружие. Поэтому, справившись с какой-либо угрозой, наши Т-лимфоциты ее запоминают. В свою очередь, организм вырабатывает специальные клетки, которые будут хранить эту информацию до новой "битвы" с данным чужеродным элементом. Этими элементами и будут Т-клетки памяти.
Вторичный агрессор (того вида, которому иммунная система уже противостояла) проникает в организм. Т-клетка памяти узнает его. Затем данная частица начинает активно размножаться, чтобы дать чужеродному организму вторичный достойный иммунный ответ.

Нормальные показатели Т-лимфоцитов в крови человека
В данной категории невозможно представить какую-то определенную цифру - нормальные значения будут варьироваться в зависимости от возраста человека. Связано это с особенностями развития его иммунной системы. С возрастом будет уменьшаться объем вилочковой железы. Поэтому, если в детстве в крови преобладают лимфоциты, то с взрослением они передают лидирующую позицию уже нейтрофилам.
Уровень Т-лимфоцитов крови помогает определить общий клинический анализ крови. Нормальные показатели тут такие:
- (50,4±3,14)*0,6-2,5 тыс.
- 50-70%.
- Соотношение "хелперы/супрессоры" - 1,5-2.

О чем говорят повышенные и пониженные показатели
Повышенное содержание Т-лимфоцитов в крови может свидетельствовать о следующем:
- Хронический или острый лимфолейкоз.
- Гиперактивный иммунитет.
- Синдром Сезари.
А, напротив, пониженное содержание Т-элементов говорит о следующих патологиях и заболеваниях:
- Хронические инфекции - гнойные процессы, ВИЧ, туберкулез.
- Сниженная выработка лимфоцитов.
- Генетические заболевания, вызывающие иммунодефицит.
- Опухоли лимфоидной ткани.
- Почечная и сердечная недостаточность, наблюдаемая на последней стадии.
- Т-клеточная лимфома.
- Прием пациентом медикаментов, которые разрушают лимфоциты.
- Следствие лучевой терапии.
Мы познакомились с Т-лимфоцитами - клетками-защитниками нашего организма. Каждый из видов выполняет при этом свою особенную функцию.
Первым исследованием всегда является подсчет лейкоцитарной формулы (см. главу «Гематологические исследования»). Оцениваются как относительные, так и абсолютные значения количества клеток периферической крови.
Определение основных популяций (Т-клетки, В-клетки, натуральные киллеры) и субпопуляций Т-лимфоцитов (Т-хелперы, Т-ЦТЛ). Для первичного исследования иммунного статуса и выявления выраженных нарушений иммунной системы ВОЗ рекомендовано определение CD3, CD4, CD8, CD19, CD16+56, соотношение CD4/CD8. Исследование позволяет определить относительное и абсолютное количество основных популяций лимфоцитов: Т-клетки – CD3, В-клетки – CD19, натуральные киллеры (NK) – CD3- CD16++56+, субпопуляции Т лимфоцитов (Т-хелперы CD3+ CD4+, Т-цитотоксические CD3+ CD8+ и их соотношение).
Метод исследования
Иммунофенотипирование лимфоцитов проводится c использованием моноклональных антител к поверхностным дифференцировочным ангинам на клетках иммунной системы, методом проточной лазерной цитофлуорометрии на проточных цитофлуориметрах.
Выбор зоны анализа лимфоцитов производится по дополнительному маркеру CD45, который представлен на поверхности всех лейкоцитов.
Условия взятия и хранения образцов
Венозная кровь, взятая из локтевой вены, утром, строго натощак, в вакуумную систему до указанной на пробирке метки. В качестве антикоагулянта используется К2ЭДТА. После взятия пробирку с образцом медленно переворачивают 8-10 раз для перемешивония крови с антикоагулянтом. Хранение и транспортировка строго при 18–23°С в вертикальном положении не более 24 ч.
Невыполнение этих условий приводит к некорректным результатам.
Интерпретация результатов
Т-лимфоциты (CD3+ клетки). Повышенное количество свидетельствует о гиперактивности иммунитета, наблюдается при острых и хронических лимфолейкозах. Увеличение относительного показателя встречается при некоторых вырусных и бактериальных инфекциях в начале заболевания, обострениях хронических заболеваний.
Снижение абсолютного количества Т-лимфоцитов свидетельствует о недостаточности клеточного иммунитета, а именно о недостаточности клеточно-эффекторного звена иммунитета. Выявляется при воспалениях разнообразной этиологии, злокачественных новообразованиях, после травмы, операций, инфаркта, при курении, приеме цитостатиков. Повышение их числа в динамике заболевания – клинически благоприятный признак.
В-лимфоциты (CD19+ клетки) Снижение наблюдается при физиологических и врожденных гипогаммаглобулинемиях и агаммаглобулинемиях, при новообразованиях иммунной системы, лечении иммунодепрессантами, острой вирусной и хронической бактериальной инфекциях, состоянии после удаления селезенки.
NK-лимфоциты с фенотипом CD3-CD16++56+ Натуральные киллеры (NK-клетки) – популяция больших гранулярных лимфоцитов. Они способны лизировать клетки-мишени, инфицированные вирусами и другими внутриклеточными антигенами, опухолевые клетки, а также другие клетки аллогенного и ксеногенного происхождения.
Увеличение количества NK-клеток связано с активацией антитрансплантационного иммунитета, в некоторых случаях отмечается при бронхиальной астме, встречается при вирусных заболеваниях, повышается при злокачественных новообразованиях и лейкозах, в периоде реконвалесценции.
Т-лимфоциты хелперы с фенотипом CD3+CD4+ Увеличение абсолютного и относительного количества наблюдается при аутоиммунных заболеваниях, может быть при аллергических реакциях, некоторых инфекционных заболеваниях. Это увеличение свидетельствует о стимуляции иммунной системы на антиген и служит подтверждением гиперреактивных синдромов.
Снижение абсолютного и относительного количества Т-клеток свидетельствует о гипореактивном синдроме с нарушением регуляторного звена иммунитета, является патогномичным признаком для ВИЧ-инфекции; встречается при хронических заболеваниях (бронхитах, пневмониях и т.д.), солидных опухолях.
Т-цитотоксические лимфоциты с фенотипом CD3+ CD8+ Повышение выявляется практически при всех хронических инфекциях, вирусных, бактериальных, протозойных инфекциях. Является характерным для ВИЧ-инфекции. Снижение наблюдается при вирусных гепатитах, герпесе, аутоиммунных заболеваниях.
Соотношение CD4+/CD8+ Исследование соотношения CD4+/CD8+ (CD3, CD4, CD8, CD4/CD8) рекомендовано только для мониторинга ВИЧ-инфекции и контроля эффективности АРВ терапии. Позволяет определить абсолютное и относительное количество Т-лимфоцитов, субпопуляций Т-хелперов, ЦТЛ и их соотношение.
Диапазон значений – 1,2–2,6. Снижение наблюдается при врожденных иммунодефицитах (синдром Ди-Джоржи, Незелофа, Вискотта-Олдрича), при вирусных и бактериальных инфекциях, хронических процессах, воздействии радиации и токсических химических веществ, множественной миеломе, стрессе, снижается с возрастом, при эндокринных заболеваниях, солидных опухолях. Является патогномичным признаком для ВИЧ-инфекции (менее 0,7).
Увеличение значения более 3 – при аутоиммунных заболеваниях, остром Т-лимфобластном лейкозе, тимоме, хроническом Т-лейкозе.
Изменение соотношения может быть связано с количеством хелперов и ЦТЛ у данного пациента. Например, снижение количества CD4+ Т-клеток при острой пневмонии в начале заболевания ведет к снижению индекса, а ЦТЛ при этом могут не измениться.
Для дополнительного исследования и выявления изменений иммунной системы при патологиях требующих оценки наличия острого или хронического воспалительного процесса и степени его активности, рекомендуется включать подсчет количества активированных Т-лимфоцитов с фенотипом CD3+HLA-DR+ и ТNK–клеток с фенотипом CD3+CD16++56+.
Т-активированные лимфоциты с фенотипом CD3+HLA-DR+ Маркер поздней активации, показатель гиперреактивности иммунитета. По экспрессии данного маркера можно судить о выраженности и силе иммунного ответа. Появляется на Т-лимфоцитах после 3-го дня острого заболевания. При благоприятном течении заболевания снижается до нормы. Увеличение экспрессии на Т-лимфоцитах может быть при многих заболеваниях, связанных с хроническим воспалением. Отмечено его повышение у пациентов с гепатитом С, пневмониями, ВИЧ-инфекцией, солидными опухолями, аутоиммунными заболеваниями.
ТNK-лимфоциты с фенотипом CD3+CD16++CD56+ Т-лимфоциты, несущие на своей поверхности маркеры CD16++ CD 56+. Эти клетки имеют свойства как Т-, так и NK-клеток. Исследование рекомендовано как дополнительный маркер при острых и хронических заболеваниях.
Снижение их в периферической крови может наблюдаться при различных органоспецифических заболеваниях и системных аутоиммунных процессах. Увеличение отмечено при воспалительных заболеваниях разной этиологии, опухолевых процессах.
Исследование ранних и поздних маркеров активации Т-лимфоцитов (CD3+CD25+, CD3-CD56+, CD95, CD8+CD38+) дополнительно назначают для оценки изменений ИС при острых и хронических заболеваниях, для диагностики, прогноза, мониторинга течения заболевания и проводимой терапии.
Т-активированные лимфоциты с фенотипом CD3+CD25+, рецeптор к ИЛ2 CD25+ – маркер ранней активации. О функциональном состоянии Т-лимфоцитов (CD3+) свидетельствует количество экспрессирующих рецепторов к ИЛ2 (CD25+). При гиперактивных синдромах количество этих клеток возрастает (острые и хронические лимфолейкозы, тимома, отторжение трансплантата), кроме того, повышение их может свидетельствовать о ранней стадии воспалительного процесса. В периферической крови их можно выявить в первые три дня болезни. Снижение числа этих клеток может наблюдаться при врожденных иммунодефицитах, аутоиммунных процессах, ВИЧ-инфекции, грибковых и бактериальных инфекциях, ионизирующей радиации, старении, отравлении тяжелыми металлами.
Т-цитотоксические лимфоциты с фенотипом CD8+CD38+ Присутствие CD38+ на ЦТЛ лимфоцитах отмечено у пациентов с разными заболеваниями. Информативный показатель при ВИЧ-инфекции, ожоговой болезни. Увеличение числа ЦТЛ с фенотипом CD8+CD38+ наблюдается при хронических воспалительных процессах, онкологических и некоторых эндокринных заболеваниях. При проведении терапии показатель снижается.
Субпопуляция натуральных киллеров с фенотипом CD3- CD56+ Молекула CD56 – адгезивная молекула, широко представленная в нервной ткани. Кроме натуральных киллеров, экспрессируется на многих типах клеток, в том число на Т-лимфоцитах.
Увеличение данного показателя свидетельствуют о расширении активности специфического клона клеток киллеров, которые имеют меньшую цитолитическую активность, чем NK-клетки с фенотипом CD3- CD16+. Количество этой популяции возрастает при гематологических опухолях (ЕК-клеточная или Т-клеточная лимфома, плазмоклеточная миелома, апластическая крупноклеточная лимфома), хронических заболеваниях, некоторых вырусных инфекциях.
Снижение отмечается при первичных иммунодефицитах, вирусных инфекциях, системных хронических заболеваниях, стрессе, лечении цитостатиками и кортикостероидами.
Рецептор CD95+ – один из рецепторов апоптоза. Апоптоз – сложный биологический процесс, необходимый для удаления из организма поврежденных, старых и инфицированных клеток. Рецептор CD95 экспрессируется на всех клетках иммунной системы. Он играет важную роль в контроле функционирования иммунной системы, так как является одним из рецепторов апоптоза. Его экспрессия на клетках определяет готовность клеток к апоптозу.
Снижение доли CD95+-лимфоцитов в крови пациентов свидетельствует о нарушении эффективности последнего этапа выбраковки дефектных и инфицированных собственных клеток, что может привести к рецидиву заболевания, хронизации патологического процесса, развитию аутоиммунных заболеваний и повышению вероятности опухолевой трансформации (к примеру, рака шейки матки при папилломотозной инфекции). Определение экспрессии CD95 имеет прогностическое значение при миело- и лимфопролифератиных заболеваниях.
Повышение интенсивности апоптоза наблюдается при вирусных заболеваниях, септических состояниях, при употреблении наркотических средств.
Активированные лимфоциты CD3+CDHLA-DR+, CD8+CD38+, CD3+CD25+, CD95. Тест отражает функциональное состояние Т-лимфоцитов и рекомендован для контроля за течением заболевания и контроля иммунотерапии при воспалительных заболеваниях разной этиологии.
Основная задача T-лимфоцитов - распознавание чужеродных или изменённых собственных антигенов в составе комплекса с молекулами MHC. Если на поверхности своих клеток будут представлены чужеродные или изменённые свои молекулы, T-лимфоцит запускает их уничтожение.
В отличие от B-лимфоцитов, T-лимфоциты не продуцируют растворимых форм антигенраспознающих молекул. Более того, большинство T-лимфоцитов не способны распознавать и связывать растворимые антигены.
Для того чтобы T-лимфоцит «обратил на антиген своё внимание», другие клетки должны каким-то образом «пропустить» антиген через себя и выставить его на своей мембране в комплексе с MHC-I или MHC-II. Это и есть феномен презентации антигена T-лимфоциту. Распознавание такого комплекса T-лимфоцитом - двойное распознавание, или MHC-рестрикция T-лимфоцитов.
АНТИГЕНРАСПОЗНАЮЩИЙ РЕЦЕПТОР T-ЛИМФОЦИТОВ
Антигенраспознающие рецепторы T-клеток - TCR состоят из цепей, принадлежащих к суперсемейству иммуноглобулинов (см. рис. 5-1). Выступающий над поверхностью клетки антигенраспознающий участок TCR - гетеродимер, т.е. состоит из двух разных полипептидных цепей. Известны два варианта TCR, обозначаемые как αβTCR и γδTCR. Эти варианты различаются составом полипептидных цепей антигенраспознающего участка. Каждый T-лимфоцит экспрессирует только 1 вариант рецептора. αβT-клетки были открыты раньше и изучены подробнее, чем γδT-лимфоциты. В связи с этим строение антигенраспознающего рецептора T-лимфоцитов удобнее описывать на примере αβTCR. Трансмембранно расположенный комплекс TCR состоит из 8 полипептидных
Рис. 6-1. Схема Т-клеточного рецептора и связанных с ним молекул
цепей (гетеродимера α- и β-цепей собственно TCR, двух вспомогательных цепей ζ, а также по одному гетеродимеру ε/δ- и ε/ γ-цепей молекулы СD3) (рис. 6-1).
. Трансмембранные цепи α и β TCR. Это 2 примерно одинаковые по размеру полипептидные цепи - α (молекулярная масса 40-60 кДа, кислый гликопротеин) и β (молекулярная масса 40-50 кДа, нейтральный или основный гликопротеин). Каждая из этих цепей содержит по 2 гликозилированных домена во внеклеточной части рецептора, гидрофобную (положительно заряженную за счёт остатков лизина и аргинина) трансмембранную часть и короткий (из 5-12 остатков аминокислот) цитоплазматический участок. Внеклеточные части обеих цепей соединены одной дисульфидной связью.
- V-область. Наружные внеклеточные (дистальные) домены обеих цепей имеют вариабельный аминокислотный состав. Они гомологичны V-области молекул иммуноглобулинов и составляют V-область TCR. Именно V-области α- и β-цепей вступают в связь с комплексом MHC-пептид.
-C-область. Проксимальные домены обеих цепей гомологичны константным областям иммуноглобулинов; это C-области TCR.
Короткий цитоплазматический участок (как α-, так и β-цепи) не может самостоятельно обеспечить проведение сигнала внутрь клетки. Для этого служат 6 дополнительных полипептидных цепей: γ, δ, 2ε и 2ζ.
.Комплекс CD3. Цепи γ, δ, ε между собой образуют гетеродимеры γε и δε (вместе их называют комплекс CD3). Этот комплекс необходим для экспрессии α- и β-цепей, их стабилизации и проведения сигнала внутрь клетки. Этот комплекс состоит из внеклеточной, трансмембранной (отрицательно заряженной и потому электростатически связанной с трансмембранными участками α- и β-цепей) и цитоплазматической частей. Важно не путать цепи CD3-комплекса с γδ-цепями димера TCR.
.ζ-Цепи соединены между собой дисульфидным мостиком. Большая часть этих цепей расположена в цитоплазме. ζ-Цепи осуществляют проведение сигнала внутрь клетки.
.ITAM-последовательности. Цитоплазматические участки полипептидных цепей γ, δ, ε и ζ содержат 10 последовательностей ITAM (1 последовательность в каждой γ-, ε- и δ-цепях и 3 - в каждой ζ-цепи), взаимодействующих с Fyn - тирозинкиназой цитозоля, активация которой инициирует начало биохимических реакций по проведению сигнала (см. рис. 6-1).
В связывании антигена участвуют ионные, водородные, ван-дерваальсовы и гидрофобные силы; конформация рецептора при этом существенно изменяется. Теоретически каждый TCR способен связывать порядка 10 5 разных антигенов, причём не только родственных по строению (перекрёстно реагирующих), но и не гомологичных по структуре. Однако в реальности полиспецифичность TCR ограничивается распознаванием всего лишь нескольких структурно схожих антигенных пептидов. Структурной основой этого феномена является особенность одновременного распознавания TCR комплекса «МНС-пептид».
Корецепторные молекулы CD4 и CD8
Помимо самого TCR каждый зрелый T-лимфоцит экспрессирует одну из так называемых корецепторных молекул - CD4 или CD8, которые также взаимодействуют с молекулами MHC на АПК или клеткахмишенях. Каждая из них имеет цитоплазматический участок, связанный
с тирозинкиназой Lck, и, вероятно, вносит свой вклад в проведение сигнала внутрь клетки при распознавании антигена.
.CD4 (β2-доменом) молекулы MHC-II (принадлежит к суперсемейству иммуноглобулинов, см. рис. 5-1, б). CD4 имеет молекулярную массу 55 кДа и 4 домена во внеклеточной части. При активации T-лимфоцита одну молекулу TCR «обслуживают» 2 молекулы CD4: вероятно, происходит димеризация молекул CD4.
.CD8 связывается с инвариантной частью (αЗ-доменом) молекулы MHC-I (принадлежит к суперсемейству иммуноглобулинов, см. рис. 5-1, а). CD8 - гетеродимер цепей α и β , соединённых дисульфидной связью. В некоторых случаях обнаруживают гомодимер из двух α-цепей, который также может взаимодействовать с MHC-I. Во внеклеточной части каждая из цепей имеет по одному иммуноглобулиноподобному домену.
Гены T-клеточного рецептора
Гены α-, β-, γ- и δ-цепей (рис. 6-2, также см. рис. 5-4) гомологичны генам иммуноглобулинов и претерпевают при дифференцировке T-лимфоцитов рекомбинацию ДНК, что теоретически обеспечивает генерацию порядка 10 16 -10 18 вариантов антигенсвязывающих рецепторов (реально это разнообразие ограничено числом лимфоцитов в организме до 10 9).
.Гены α-цепи имеют ~54 V-сегмента, 61 J- и 1 C-сегмент.
.Гены β-цепи содержат ~65 V-сегментов, 2 D-сегмента, 13 J-сегментов и 2 C-сегмента.
.Гены δ-цепи. Между V- и J-сегментами α-цепи расположены гены D-(3), J-(4) и C-(1) сегментов δ-цепи γδTCR. V-сегменты δ-цепи «вкраплены» среди V-сегментов α-цепи.
.Гены γ-цепи γδTCR имеют 2 C-сегмента, 3 J-сегмента перед первым C-сегментом и 2 J-сегмента перед вторым C-сегментом, 15 V-сегментов.
Перестройка генов
.Рекомбинация ДНК происходит при объединении V-, D- и J-сегментов и катализируется тем же комплексом рекомбиназ, что и при дифференцировке B-лимфоцитов.
.После перестройки VJ в генах α-цепи и VDJ в генах β-цепи, а также после присоединения некодируемых N- и P-нуклеотидов с ДНК

Рис. 6-2. Гены α- и β-цепей антигенраспознающего рецептора T-лимфоцитов человека
транскрибируется РНК. Объединение с C-сегментом и удаление лишних (неиспользуемых) J-сегментов происходит при сплайсинге первичного транскрипта.
. Гены α-цепи могут перестраиваться неоднократно при уже правильно перестроенных и экспрессированных генах β-цепи. Именно поэтому есть некоторая вероятность того, что одна клетка может нести более одного варианта TCR.
. Соматическому гипермутагенезу гены TCR не подвергаются.
ПРОВЕДЕНИЕ СИГНАЛА С АНТИГЕНРАСПОЗНАЮЩИХ РЕЦЕПТОРОВ ЛИМФОЦИТОВ
TCR и BCR имеют ряд общих закономерностей регистрации и проведения в клетку активационных сигналов (см. рис. 5-11).
. Кластеризация рецепторов. Для активации лимфоцита необходима кластеризация антигенраспознающих рецепторов и корецепторов, т.е. «сшивка» нескольких рецепторов одним антигеном.
. Тирозинкиназы. В проведении сигнала играют значительную роль процессы фосфорилирования/дефосфорилирования белков по остатку тирозина под действием тирозинкиназ и тирозинфосфатаз,
ведущие к активации или инактивации этих белков. Эти процессы легко обратимы и «удобны» для быстрых и гибких реакций клетки на внешние сигналы.
. Киназы Src. Богатые тирозином ITAM-последовательности цитоплазматических участков иммунорецепторов подвергаются фосфорилированию под действием нерецепторных (цитоплазматических) тирозинкиназ семейства Src (Fyn, Blk, Lyn в B-лимфоцитах, Lck и Fyn - в T-лимфоцитах).
. Киназы ZAP-70 (в T-лимфоцитах) или Syk (в B-лимфоцитах), связываясь с фосфорилированными ITAM-последовательностями, активируются и начинают фосфорилировать адапторные белки: LAT (Linker for Activation of T cells) (киназой ZAP-70), SLP-76 (киназой ZAP-70) или SLP-65 (киназой Syk).
. Адапторные белки рекрутируют фосфоинозитид-3-киназу (PI3K). Эта киназа в свою очередь активирует серин/треониновую протеинкиназу Akt, вызывая усиление белкового биосинтеза, что способствует ускоренному росту клеток.
. Фосфолипаза C γ(см. рис. 4-8). Киназы семейства Tec (Btk - в B-лимфоцитах, Itk - в T-лимфоцитах) связывают адапторные белки и активируют фосфолипазу Cγ(PLCγ).
PLCγрасщепляет фосфатидилинозитдифосфат (PIP 2) клеточной мембраны на инозит-1,4,5-трифосфат (IP 3) и диацилглицерин
(DAG).
DAG остаётся в мембране и активирует протеинкиназу С (PKC) - серин/треониновую киназу, которая активирует эволюционно «древний» фактор транскрипции NFκB.
IP 3 связывается со своим рецептором в эндоплазматическом ретикулуме и высвобождает ионы кальция из депо в цитозоль.
Свободный кальций активирует кальцийсвязывающие белки - кальмодулин, регулирующий активность ряда других белков, и кальциневрин, дефосфорилирующий и тем самым активирующий ядерный фактор активированных T-лимфоцитов NFAT (Nuclear Factor of Activated T cells).
. Ras и другие малые G-белки в неактивном состоянии связаны с ГДФ, но адапторные белки заменяют последний на ГТФ, чем переводят Ras в активное состояние.
Ras обладает собственной ГТФазной активностью и быстро отщепляет третий фосфат, чем возвращает себя в неактивное состояние (самоинактивируется).
В состоянии кратковременной активации Ras успевает активировать очередной каскад киназ, называемых MAPK (MitogenActivated Protein Kinase), которые в итоге активируют фактор транскрипции AP-1 в ядре клетки. На рис. 6-3 схематично представлены основные пути передачи сигналов с TCR. Активационный сигнал включается при связывании TCR с лигандом (комплексом молекула МНС-пептид) при участии корецептора (CD4 или CD8) и костимулирующей молекулы CD28. Это приводит к активации киназ Fyn и Lck. Красным цветом отмечены участки ITAM в цитоплазматических частях полипептидных цепей CD3. Отражена роль Src-киназ, связанных с рецептором, в фосфорилировании белков: как рецепторных, так и сигнальных. Обращает на себя внимание чрезвычайно широкий спектр эффектов киназы Lck, связанной с корецепторами; роль киназы Fyn установлена с меньшей определённостью (отражено в прерывистом характере линий).

Рис. 6-3. Источники и направление пусковых активационных сигналов при стимуляции Т-лимфоцитов. Обозначения: ZAP-70 (ζ-associated proteinkinase, мол. масса 70 кДа) - протеинкиназа р70, связанная с ζ-цепью; PLCγ (Phospholipase С γ) - фосфолипаза С, изоформа γ; PI3K (Phosphatidyl Inositol 3-kinase) - фосфатидилинозитол 3-киназа; Lck, Fyn -тирозинкиназы; LAT, Grb, SLP, GADD, Vav - адапторные белки
Ключевую роль в посредничестве между рецепторными киназами и адапторными молекулами и ферментами играет тирозинкиназа ZAP-70. Она активирует (через фосфорилирование) адапторные молекулы SLP-76 и LAT, а последняя передаёт активационный сигнал другим адапторным белкам GADD, GRB и активирует у-изоформу фосфолипазы С (PLCy). До этого этапа в передачу сигнала вовлекаются исключительно факторы, связанные с клеточной мембраной. Важный вклад во включение сигнальных путей вносит костимулирующая молекула CD28, реализующая своё действие через связанную с ней липидную киназу PI3K (Phosphatidyl Inositol 3-kinase). Основной мишенью киназы PI3K служит фактор Vav, связанный с цитоскелетом.
В результате формирования сигнала и передачи его от рецептора Т-клетки к ядру образуются 3 транскрипционных фактора - NFAT, AP-1 и NF-kB, индуцирующие экспрессию генов, контролирующих процесс активации Т-лимфоцитов (рис. 6-4). К образованию NFAT приводит сигнальный путь, не зависящий от костимуляции, который включается благодаря активации фосфолипазы С и реализуется с участием ионов

Рис. 6-4. Схема сигнальных путей при активации Т-клеток. NFAT (Nuclear factor of activated T cells), AP-1 (Activation protein-1), NF-κB (Nuclear factor of к -gene of B cells) - факторы транскрипции
Са 2+ . Этот путь вызывает активацию кальциневрина, который, обладая активностью фосфатазы, дефосфорилирует цитозольный фактор NFAT-Р. Благодаря этому NFAT-Р приобретает способность мигрировать в ядро и связываться с промоторами активационных генов. Фактор АР-1 формируется как гетеродимер из белков с-Fos и с-Jun, образование которых индуцируется благодаря активации соответствующих генов под влиянием факторов, образующихся в результате реализации трёх компонентов МАР-каскада. Эти пути включаются при участии коротких ГТФ-связывающих белков Ras и Rac. Значительный вклад в реализацию МАР-каскада вносят сигналы, зависящие от костимуляции через молекулу CD28. Третий транскрипционный фактор, NF-kB, известен как основной транскрипционный фактор клеток врождённого иммунитета. Он активируется в результате расщепления блокирующей субъединицы IkB киназой IKK, которая в Т-клетках активируется в ходе передачи сигнала, зависимого от изоформы ϴ протеинкиназы С (PKC9). Основной вклад во включение этого сигнального пути вносят костимулирующие сигналы от CD28. Сформировавшиеся транскрипционные факторы, связавшись с промоторными участками генов, индуцируют их экспрессию. Для начальных этапов реакции Т-клеток на стимуляцию особенно важна экспрессия генов IL2 и IL2R, что обусловливает выработку ростового фактора Т-клеток ИЛ-2 и экспрессию его высокоаффинного рецептора на Т-лимфоцитах. В результате ИЛ-2 выступает как аутокринный ростовой фактор, обусловливающий пролиферативную экспансию Т-клеток клонов, вовлечённых в реакцию на антиген.
ДИФФЕРЕНЦИРОВКА T-ЛИМФОЦИТОВ
В основе выделения этапов развития Т-лимфоцитов лежит состояние рецепторных V-генов и экспрессии TCR, а также корецепторов и других мембранных молекул. Схема дифференцировки Т-лимфоцитов (рис. 6-5) аналогична приведённой выше схеме развития В-лимфоцитов (см. рис. 5-13). Приведены ключевые характеристики фенотипа и ростовых факторов развивающихся Т-клеток. Принятые обозначения стадий развития Т-клеток определяются экспрессией корецепторов: DN (от Double-Negative, CD4CD8) - двойные отрицательные, DP (от Double-Positive, CD4 + CD8 +) - двойные положительные, SP (от Single-Positive, CD4 + CD8 - и CD4CD8 +) - одинарно положительные. Деление DNтимоцитов на стадии DN1, DN2, DN3 и DN4 основывается на характере

Рис. 6-5. Развитие Т-лимфоцитов
экспрессии молекул CD44 и CD25. Другие условные обозначения: SCF (от Stem Cell Factor) - фактор стволовых клеток, lo (low; метка индекса) - низкий уровень экспрессии. Стадии реаранжировки: D-J - предварительный этап, соединение сегментов D и J (только в генах β- и δ-цепей TCR, см. рис. 6-2), V-DJ - завершающий этап, соединение зародышевого V-гена с объединённым сегментом DJ.
.Тимоциты дифференцируются из общей клетки-предшественника, которая ещё вне тимуса экспрессирует такие мембранные маркёры, как CD7, CD2, CD34 и цитоплазматическую форму CD3.
.Коммитированные к дифференцировке в T-лимфоциты клеткипредшественники мигрируют из костного мозга в субкапсулярную зону коры тимуса, где примерно в течение одной недели медленно пролиферируют. На тимоцитах появляются новые мембранные молекулы CD44 и CD25.
.Затем клетки перемещаются вглубь коры тимуса, молекулы CD44 и CD25 исчезают с их мембраны. В этой стадии начинается перестройка генов β -, γ- и δ-цепей TCR. Если гены γ- и δ-цепей успевают продуктивно, т.е. без сдвига рамки считывания, перестроиться раньше, чем гены β-цепи, то лимфоцит дифференцируется далее как γδT. В противном случае происходит экспрессия β-цепи на мембране в комплексе с pT α (инвариантной суррогатной цепью, заменяющей на этом этапе настоящую α-цепь) и CD3. Это служит
сигналом к прекращению перестройки генов γ- и δ-цепей. Клетки начинают пролиферировать и экспрессировать одновременно CD4 и CD8 - дважды позитивные тимоциты. При этом накапливается масса клеток с уже готовой β-цепью, но с ещё не перестроенными генами α-цепи, что вносит свой вклад в разнообразие αβ-гетеродимеров.
.На следующем этапе клетки перестают делиться и начинают перестраивать Vα-гены, причём несколько раз в течение 3-4 сут. Перестройка генов α-цепи приводит к необратимой делеции δ-локуса, расположенного между сегментами генов α-цепи.
.Происходят экспрессия TCR с каждым новым вариантом α-цепи и отбор (селекция) тимоцитов по силе связывания с комплексом MHC-пептид на мембранах эпителиальных клеток тимуса.
Позитивная селекция: погибают тимоциты, не связавшие ни одного из доступных комплексов MHC-пептид. В результате позитивной селекции в тимусе погибает около 90% тимоцитов.
Негативная селекция уничтожает клоны тимоцитов, связывающих комплексы MHC-пептид со слишком высокой аффинностью. Негативная селекция элиминирует от 10 до 70% клеток, прошедших позитивную селекцию.
Тимоциты, связавшие какой-либо из комплексов MHC-пептид с правильной, т.е. средней по силе, аффинностью, получают сигнал к выживанию и продолжают дифференцировку.
.На короткое время с мембраны тимоцитов исчезают обе корецепторные молекулы, а затем экспрессируется одна из них: тимоциты, распознавшие пептид в комплексе с MHC-I, экспрессируют корецептор CD8, а с MHC-II - корецептор CD4. Соответственно на периферию выходят (в соотношении около 2:1) T-лимфоциты двух типов: CD8 + и CD4 + , функции которых в предстоящих иммунных ответах различны.
-CD8 + T-клетки играют роль цитотоксических T-лимфоцитов (ЦТЛ) - они распознают и непосредственно убивают клетки, модифицированные вирусом, опухолевые и другие «изменённые» клетки (рис. 6-6).
-CD4 + T-клетки. Функциональная специализация CD4 + T-лимфоцитов более разнообразна. Значительная часть CD4 + T-лимфоцитов в процессе развития иммунного ответа становится T-хелперами (помощниками), взаимодействующими с В-лимфоцитами, Т-лимфоцитами и другими клетками при

Рис. 6-6. Механизм воздействия цитотоксического T-лимфоцита на клеткумишень. В Т-киллере в ответ на увеличение концентрации Са 2+ гранулы с перфорином (фиолетовые овалы) и гранзимами (жёлтые кружочки) сливаются с клеточной мембраной. Освободившийся перфорин встраивается в мембрану клетки-мишени с последующим образованием пор, проницаемых для гранзимов, воды и ионов. В результате клетка-мишень лизируется
прямом контакте или через растворимые факторы (цитокины). В определённых случаях из них могут развиться CD4 + ЦТЛ: в частности, такие T-лимфоциты обнаружены в значительных количествах в коже больных с синдромом Лайелла.
Субпопуляции T-хелперов
С конца 80-х годов XX века было принято выделять 2 субпопуляции T-хелперов (в зависимости от того, какой набор цитокинов они продуцируют) - Th1 и Th2. В последние годы спектр субпопуляций CD4 + Т-клеток продолжает расширяться. Обнаружены такие субпопуляции, как: Th17, T-регуляторы, Tr1, Th3, Tfh и др.
Основные субпопуляции CD4 + Т-клеток:
. Th0 - CD4 + Т-лимфоциты на ранних стадиях развития иммунного ответа, они продуцируют только ИЛ-2 (митоген для всех лимфоцитов).
.Th1 - дифференцированная субпопуляция CD4 + Т-лимфоцитов, специализирующаяся на продукции ИФН γ, ФНО β и ИЛ-2. Эта субпопуляция осуществляет регуляцию многих реакций клеточного иммунитета, включая гиперчувствительность замедленного типа (ГЗТ) и активацию ЦТЛ. Кроме того, Th1 стимулируют продукцию В-лимфоцитами опсонизирующих антител класса IgG, запускающих каскад активации комплемента. Развитие избыточного воспаления с последующим повреждением тканей напрямую связано с активностью Th1-субпопуляции.
.Th2 - дифференцированная субпопуляция CD4 + Т-лимфоцитов, специализирующаяся на выработке ИЛ-4, ИЛ-5, ИЛ-6, ИЛ-10 и ИЛ-13. Эта субпопуляция участвует в активации В-лимфоцитов и способствует секреции ими больших количеств антител разных классов, особенно IgE. Кроме того, Th2-субпопуляция участвует в активации эозинофилов и развитии аллергических реакций.
.Th17 - субпопуляция CD4 + Т-лимфоцитов, специализирующаяся на образовании ИЛ-17. Эти клетки осуществляют противогрибковую и антимикробную защиту эпителиальных и слизистых барьеров, а также играют ключевую роль в патологии аутоиммунных заболеваний.
.Т-регуляторы - CD4 + Т-лимфоциты, подавляющие активность других клеток иммунной системы посредством секреции иммуносупрессорных цитокинов - ИЛ-10 (ингибитора активности макрофагов и Th1-клеток) и ТФРβ - ингибитора пролиферации лимфоцитов. Ингибиторный эффект может также достигаться при непосредственном межклеточном взаимодействии, поскольку на мембране некоторых Т-регуляторов экспрессированы индукторы апоптоза активированных и «отработавших» лимфоцитов - FasL (Fas-лиганд). Существует несколько популяций CD4 + регуляторных Т-лимфоцитов: естественные (Treg), созревающие в тимусе (CD4 + CD25 + , экспрессируют фактор транскрипции Foxp3), и индуцированные - локализованные преимущественно в слизистых оболочках пищеварительного тракта и переключившиеся на образование ТФРβ (Th3) или ИЛ-10 (Tr1). Нормальное функционирование Т-регуляторов необходимо для поддержания гомеостаза иммунной системы и предотвращения развития аутоиммунных заболеваний.
.Дополнительные хелперные популяции. В последнее время появляется описание всё новых популяций CD4 + Т-лимфоцитов, клас-
сифицированных по типу преимущественно продуцируемого ими цитокина. Так, как оказалось, одной из важнейших популяций являются Tfh (от англ. follicular helper - фолликулярный хелпер). Эта популяция CD4 + Т-лимфоцитов преимущественно расположена в лимфоидных фолликулах и осуществляет хелперную функцию для В-лимфоцитов посредством продукции ИЛ-21, вызывая их созревание и терминальную дифференцировку в плазматические клетки. Кроме ИЛ-21 Tfh могут также продуцировать ИЛ-6 и ИЛ-10, необходимые для дифференцировки В-лимфоцитов. Нарушение функций этой популяции приводит к развитию аутоиммунных заболеваний или иммунодефицитов. Другой «новоявленной» популяцией являются Th9 - продуценты ИЛ-9. По-видимому, это Th2, переключившиеся на секрецию ИЛ-9, способного вызывать пролиферацию Т-хелперных клеток при отсутствии антигенной стимуляции, а также усиливать секрецию В-лимфоцитами IgM, IgG и IgE.
Основные субпопуляции Т-хелперов представлены на рис. 6-7. На рисунке суммированы современные представления об адаптивных субпопуляциях CD4 + Т-клеток, т.е. субпопуляций, формирующих-

Рис. 6-7. Адаптивные субпопуляции CD4 + Т-клеток (цитокины, дифференцировочные факторы, хемокиновые рецепторы)
ся при иммунном ответе, а не в ходе естественного развития клеток. Для всех разновидностей Т-хелперов указаны цитокины-индукторы (на стрелках, ведущих к кружкам, символизирующим клетки), транскрипционные факторы (внутри кружков), хемокиновые рецепторы, направляющие миграцию (около линий, отходящих от «поверхности клетки»), и продуцируемые цитокины (в прямоугольниках, на которые направлены стрелки, отходящие от кружков).
Расширение семейства адаптивных субпопуляций CD4 + Т-клеток потребовало решения вопроса о природе клеток, с которыми взаимодействуют эти субпопуляции (кому они оказывают «помощь» в соответствии со своей функцией хелперов). Эти представления отражены на рис. 6-8. Здесь же представлен уточнённый взгляд на функции этих субпопуляций (участие в защите от определённых групп патогенов), а также о патологических последствиях несбалансированного усиления активности этих клеток.

Рис. 6-8. Адаптивные субпопуляции Т-клеток (клетки-партнёры, физиологические и патологические эффекты)
γ δT-лимфоциты
Подавляющее большинство (99%) T-лимфоцитов, проходящих лимфопоэз в тимусе, составляют αβT-клетки; менее 1% - γδT-клетки. Последние в большинстве дифференцируются вне тимуса, в первую очередь в слизистых оболочках пищеварительного тракта. В коже, лёгких, пищеварительном и репродуктивном трактах они являются доминирующей субпопуляцией внутриэпителиальных лимфоцитов. Среди всех T-лимфоцитов организма γδT-клетки составляют от 10 до 50%. В эмбриогенезе γδT-клетки появляются раньше αβT-клеток.
.γδT-клетки не экспрессируют CD4. Молекула CD8 экспрессирована на части γδT-клеток, но не в виде ap-гетеродимера, как на CD8 + apT-клетках, а в виде гомодимера из двух a-цепей.
.Антигенраспознающие свойства: γδTCR в большей степени напоминают иммуноглобулины, чем αβTCR, т.е. способны связывать нативные антигены независимо от классических молекул MHC - для γδT-клеток не обязателен или вовсе не нужен предварительный процессинг антигена АПК.
.Разнообразие γδTCR меньше, чем αβTCR или иммуноглобулинов, хотя в целом γδT-клетки способны распознавать широкий спектр антигенов (в основном это фосфолипидные антигены микобактерий, углеводы, белки теплового шока).
.Функции γδT-клеток ещё до конца не изучены, хотя становится преобладающим мнение, что они служат одним из связующих компонентов между врождённым и приобретённым иммунитетом. γδT-клетки - один из первых барьеров на пути патогенов. Кроме того, эти клетки, секретируя цитокины, играют важную иммунорегуляторную роль и способны дифференцироваться в ЦТЛ.
NKT-лимфоциты
Естественные киллерные Т-клетки (NKT-клетки) представляют особую субпопуляцию лимфоцитов, занимающую промежуточное положение между клетками врождённого и адаптивного иммунитета. Эти клетки имеют черты как NK-, так и Т-лимфоцитов. NKT-клетки экспрессируют αβTCR и характерный для NK-клеток рецептор NK1.1, принадлежащий к суперсемье лектиновых гликопротеинов С-типа. Однако TCR-рецептор NKT-клеток имеет существенные отличия от TCR-рецептора обычных клеток. У мышей большинство NKTклеток экспрессирует инвариантный V-домен a-цепи, состоящий из
сегментов Vα14-Jα18, иногда обозначаемый как Jα281. У человека V-домен α-цепи состоит из сегментов Vα24-JαQ. У мышей α-цепь инвариантного TCR преимущественно комплексируется с Vβ8.2, у человека - с Vβ11. Из-за особенностей строения цепей TCR NKTклеток называют инвариантным - iTCR. Развитие NKT-клеток зависит от молекулы CD1d, которая имеет сходство с молекулами МНС-I. В отличие от классических молекул МНС-I, презентирующих Т-клеткам пептиды, CD1d презентирует Т-клеткам только гликолипиды. Хотя считается, что печень является местом развития NKT-клеток, имеются строгие доказательства роли тимуса в их развитии. NKT-клетки играют важную роль в регуляции иммунитета. У мышей и людей с различными аутоиммунными процессами функциональная активность NKT-клеток сильно нарушена. Полной картины значимости таких нарушений в патогенезе аутоиммунных процессов нет. При некоторых аутоиммунных процессах NKT-клетки могут играть супрессорную роль.
Помимо контроля аутоиммунных и аллергических реакций, NKTклетки участвуют в иммунном надзоре, вызывая при повышении функциональной активности отторжение опухолей. Велика их роль в противомикробной защите, особенно на ранних этапах развития инфекционного процесса. NKT-клетки вовлекаются в различные воспалительные инфекционные процессы, особенно при вирусных поражениях печени. В целом NKT-клетки - многофункциональная популяция лимфоцитов, несущая ещё много научных загадок.
На рис. 6-9 обобщены данные о дифференцировке Т-лимфоцитов на функциональные субпопуляции. Представлены несколько уровней бифуркации: γ δТ/ αβТ, далее для αβТ-клеток - NKT/ остальные Т-лимфоциты, для последних - CD4 + /CD8 + , для CD4 + Т-клеток - Th/Treg, для CD8 + Т-лимфоцитов - CD8αβ/CD8αα. Показаны также дифференцировочные транскрипционные факторы, ответственные за все линии развития.

Рис. 6-9. Естественные субпопуляции Т-лимфоцитов и их дифференцировочные факторы